 Ulaz
UlazInterakcija ćelije sa okolinom. Odnos organizma i okoline Razvoj zametnih ćelija u biljkama i životinjama
Pozivamo vas da se upoznate sa materijalima i.
: celulozna membrana, membrana, citoplazma sa organelama, jezgro, vakuole sa ćelijskim sokom.Prisustvo plastida je glavna karakteristika biljne ćelije.
Funkcije ćelijske membrane- određuje oblik ćelije, štiti od faktora okoline.
Plazma membrana- tanak film, koji se sastoji od interakcijskih molekula lipida i proteina, omeđuje unutrašnji sadržaj od vanjskog okruženja, osigurava transport vode, minerala i organskih tvari u ćeliju putem osmoze i aktivnog transporta, a također uklanja otpadne produkte.
Citoplazma- unutrašnje polutečno okruženje ćelije, u kojem se nalaze jezgro i organele, obezbeđuje veze između njih i učestvuje u osnovnim životnim procesima.
Endoplazmatski retikulum- mreža razgranatih kanala u citoplazmi. Učestvuje u sintezi proteina, lipida i ugljikohidrata, te u transportu tvari. Ribosomi su tijela smještena na ER ili u citoplazmi, koja se sastoje od RNK i proteina, a uključena su u sintezu proteina. EPS i ribozomi su jedan aparat za sintezu i transport proteina.
Mitohondrije- organele odvojene od citoplazme sa dvije membrane. U njima se oksidiraju organske tvari i sintetiziraju se molekule ATP-a uz sudjelovanje enzima. Povećanje površine unutrašnje membrane na kojoj se nalaze enzimi zbog krista. ATP je organska supstanca bogata energijom.
Plastidi(hloroplasti, leukoplasti, hromoplasti), njihov sadržaj u ćeliji je glavna karakteristika biljnog organizma. Kloroplasti su plastidi koji sadrže zeleni pigment hlorofil, koji apsorbira svjetlosnu energiju i koristi je za sintezu organskih tvari iz ugljičnog dioksida i vode. Kloroplasti su odvojeni od citoplazme sa dvije membrane, brojnim izraslinama - grana na unutrašnjoj membrani, u kojoj se nalaze molekule hlorofila i enzimi.
Golgijev kompleks- sistem šupljina ograničenih od citoplazme membranom. Akumulacija proteina, masti i ugljikohidrata u njima. Provođenje sinteze masti i ugljikohidrata na membranama.
Lizozomi- tijela odvojena od citoplazme jednom membranom. Enzimi koje sadrže ubrzavaju razgradnju složenih molekula na jednostavne: proteina u aminokiseline, složenih ugljikohidrata u jednostavne, lipida u glicerol i masne kiseline, a uništavaju i mrtve dijelove stanice i cijele stanice.
Vakuole- šupljine u citoplazmi ispunjene ćelijskim sokom, mjesto nakupljanja rezervnih hranjivih i štetnih materija; regulišu sadržaj vode u ćeliji.
Core- glavni dio ćelije, sa vanjske strane prekriven dvomembranskim nuklearnim omotačem probušenim porama. Supstance ulaze u jezgro i uklanjaju se iz njega kroz pore. Kromosomi su nosioci nasljednih informacija o karakteristikama organizma, glavnim strukturama jezgra, od kojih se svaka sastoji od jednog molekula DNK u kombinaciji s proteinima. Jezgro je mjesto sinteze DNK, mRNA i rRNA.

Prisustvo vanjske membrane, citoplazme sa organelama i jezgra s hromozomima.
Vanjska ili plazma membrana- omeđuje sadržaj ćelije od okoline (ostale ćelije, međućelijska supstanca), sastoji se od molekula lipida i proteina, obezbeđuje komunikaciju između ćelija, transport supstanci u ćeliju (pinocitoza, fagocitoza) i van ćelije.
Citoplazma- unutrašnje polutečno okruženje ćelije, koje obezbeđuje komunikaciju između jezgra i organela koji se nalaze u njoj. Glavni životni procesi odvijaju se u citoplazmi.
ćelijske organele:
1) endoplazmatski retikulum (ER)- sistem granajućih tubula, učestvuje u sintezi proteina, lipida i ugljenih hidrata, u transportu supstanci u ćeliji;
2) ribozomi- tijela koja sadrže rRNA nalaze se na ER i u citoplazmi i učestvuju u sintezi proteina. EPS i ribozomi su jedan aparat za sintezu i transport proteina;
3) mitohondrije- "elektrane" ćelije, odvojene od citoplazme sa dve membrane. Unutrašnji formira kriste (nabore), povećavajući svoju površinu. Enzimi na kristama ubrzavaju oksidaciju organskih supstanci i sintezu energetski bogatih ATP molekula;
4) Golgijev kompleks- grupa šupljina ograničenih membranom od citoplazme, ispunjenih proteinima, mastima i ugljikohidratima, koji se ili koriste u vitalnim procesima ili uklanjaju iz ćelije. Membrane kompleksa vrše sintezu masti i ugljikohidrata;
5) lizozomi- tijela ispunjena enzimima ubrzavaju razgradnju proteina na aminokiseline, lipida u glicerol i masne kiseline, polisaharida u monosaharide. U lizosomima se uništavaju mrtvi dijelovi ćelije, cijele ćelije.
Ćelijske inkluzije- akumulacije rezervnih nutrijenata: proteina, masti i ugljenih hidrata.
Core- najvažniji deo ćelije. Prekriven je dvostrukom membranom s porama, kroz koje neke tvari prodiru u jezgro, a druge ulaze u citoplazmu. Hromozomi su glavne strukture jezgra, nosioci nasljednih informacija o karakteristikama organizma. Prenosi se tokom diobe matične ćelije na ćerke ćelije, a sa zametnim ćelijama na ćerke organizme. Jezgro je mjesto sinteze DNK, mRNA i rRNA.
vježba:
Objasnite zašto se organele nazivaju specijalizirane ćelijske strukture?
odgovor: Organele se nazivaju specijalizirane ćelijske strukture, budući da obavljaju strogo određene funkcije, nasljedne informacije se pohranjuju u jezgri, ATP se sintetizira u mitohondrijima, fotosinteza se događa u hloroplastima itd.
Ako imate pitanja o citologiji, možete se obratiti
Teorija za zadatak 5 sa Jedinstvenog državnog ispita iz biologije
Struktura ćelije. Odnos između strukture i funkcija dijelova i organela ćelije je osnova njenog integriteta
Struktura ćelije
Struktura prokariotskih i eukariotskih ćelija
Glavne strukturne komponente ćelija su plazma membrana, citoplazma i nasljedni aparat. Ovisno o karakteristikama organizacije razlikuju se dvije glavne vrste ćelija: prokariotske i eukariotske. Glavna razlika između prokariotskih i eukariotskih ćelija je organizacija njihovog nasljednog aparata: kod prokariota se nalazi direktno u citoplazmi (ovo područje citoplazme se naziva nukleoid) i nije odvojen od njega membranskim strukturama, dok je kod eukariota većina DNK koncentrisana u jezgru, okružena dvostrukom membranom. Osim toga, genetske informacije prokariotskih ćelija, koje se nalaze u nukleoidu, zapisane su u kružnom DNK molekulu, dok su kod eukariota molekule DNK otvorene.
Za razliku od eukariota, citoplazma prokariotskih stanica također sadrži mali broj organela, dok eukariotske stanice karakterizira značajna raznolikost ovih struktura.
Struktura i funkcije bioloških membrana
Struktura biomembrane. Membrane koje ograničavaju ćelije i membranske organele eukariotskih ćelija imaju zajednički hemijski sastav i strukturu. Oni uključuju lipide, proteine i ugljikohidrate. Membranski lipidi su uglavnom predstavljeni fosfolipidima i holesterolom. Većina membranskih proteina su složeni proteini, kao što su glikoproteini. Ugljikohidrati se ne pojavljuju samostalno u membrani, oni su povezani s proteinima i lipidima. Debljina membrane je 7-10 nm.
Prema trenutno opšteprihvaćenom modelu fluidnog mozaika strukture membrane, lipidi formiraju dvostruki sloj, tj. lipidni dvosloj, u kojem su hidrofilne "glave" molekula lipida okrenute prema van, a hidrofobni "repovi" su skriveni unutar membrane. Ovi “repovi” zbog svoje hidrofobnosti osiguravaju odvajanje vodenih faza unutrašnjeg okruženja ćelije i njenog okruženja. Proteini su povezani s lipidima kroz različite vrste interakcija. Neki proteini se nalaze na površini membrane. Takvi proteini se nazivaju periferni, ili površno. Ostali proteini su djelomično ili potpuno uronjeni u membranu - to su integralni, ili potopljeni proteini. Membranski proteini obavljaju strukturnu, transportnu, katalitičku, receptorsku i druge funkcije.
Membrane nisu poput kristala, njihove komponente su stalno u pokretu, zbog čega se pojavljuju praznine između molekula lipida - pora kroz koje različite tvari mogu ući ili izaći iz stanice.
Biološke membrane se razlikuju po svom položaju u ćeliji, hemijskom sastavu i funkcijama. Glavne vrste membrana su plazma i unutrašnje. Plazma membrana sadrži oko 45% lipida (uključujući glikolipide), 50% proteina i 5% ugljikohidrata. Lanci ugljikohidrata, koji su dio složenih proteina-glikoproteina i složenih lipida-glikolipida, strše iznad površine membrane. Glikoproteini plazmaleme su izuzetno specifični. Na primjer, koriste se za međusobno prepoznavanje stanica, uključujući spermu i jaje.
Na površini životinjskih ćelija, ugljikohidratni lanci formiraju tanak površinski sloj - glikokaliks. Otkriva se u skoro svim životinjskim ćelijama, ali stepen njegove ekspresije varira (10-50 µm). Glikokaliks obezbeđuje direktnu komunikaciju između ćelije i spoljašnjeg okruženja, gde se dešava ekstracelularna probava; Receptori se nalaze u glikokaliksu. Pored plazmaleme, ćelije bakterija, biljaka i gljiva su takođe okružene ćelijskim membranama.
Unutrašnje membrane eukariotske ćelije razgraničavaju različite dijelove ćelije, formirajući neobične "kompartmente" - pretinci, koji pospješuje razdvajanje različitih metaboličkih i energetskih procesa. Mogu se razlikovati po hemijskom sastavu i funkcijama, ali njihov opšti strukturni plan ostaje isti.
Funkcije membrane:
- Ograničavanje. Ideja je da odvajaju unutrašnji prostor ćelije od spoljašnjeg okruženja. Membrana je polupropusna, odnosno kroz nju mogu slobodno proći samo one tvari koje su potrebne ćeliji, a postoje i mehanizmi za transport potrebnih tvari.
- Receptor. On je prvenstveno povezan sa percepcijom signala okoline i prenosom ovih informacija u ćeliju. Za ovu funkciju odgovorni su posebni proteini receptora. Membranski proteini su također odgovorni za ćelijsko prepoznavanje po principu „prijatelj ili neprijatelj“, kao i za formiranje međućelijskih veza, od kojih su najviše proučavane sinapse nervnih ćelija.
- Katalitički. Na membranama se nalaze brojni enzimski kompleksi, zbog čega se na njima odvijaju intenzivni sintetički procesi.
- Energetska transformacija. Povezan sa stvaranjem energije, njenim skladištenjem u obliku ATP-a i potrošnjom.
- Kompartmentalizacija. Membrane također ograničavaju prostor unutar ćelije, odvajajući tako početne materijale reakcije i enzime koji mogu izvršiti odgovarajuće reakcije.
- Formiranje međućelijskih kontakata. Unatoč činjenici da je debljina membrane toliko mala da se ne može razlikovati golim okom, ona, s jedne strane, služi kao prilično pouzdana barijera za ione i molekule, posebno one topive u vodi, a s druge strane , osigurava njihov transport u i iz ćelije.
- Transport.

Membranski transport. Zbog činjenice da su ćelije, kao elementarni biološki sistemi, otvoreni sistemi, za obezbeđivanje metabolizma i energije, održavanje homeostaze, rasta, razdražljivosti i drugih procesa, potreban je prenos supstanci kroz membranu - membranski transport. Trenutno se transport tvari kroz ćelijsku membranu dijeli na aktivnu, pasivnu, endo- i egzocitozu.
Pasivni transport- Ovo je vid transporta koji se odvija bez potrošnje energije od viših do nižih koncentracija. Male nepolarne molekule rastvorljive u lipidima (O 2, CO 2) lako prodiru u ćeliju jednostavna difuzija. One netopive u lipidima, uključujući nabijene male čestice, pokupe se proteini nosači ili prolaze kroz posebne kanale (glukoza, aminokiseline, K+, PO 4 3-). Ova vrsta pasivnog transporta se naziva olakšanu difuziju. Voda ulazi u ćeliju kroz pore u lipidnoj fazi, kao i kroz posebne kanale obložene proteinima. Transport vode kroz membranu naziva se osmozom.

Osmoza je izuzetno važna u životu ćelije, jer ako se stavi u rastvor sa većom koncentracijom soli nego u ćelijskom rastvoru, tada će voda početi da napušta ćeliju i volumen živog sadržaja počinje da se smanjuje. U životinjskim ćelijama ćelija se u celini smanjuje, a u biljnim ćelijama citoplazma zaostaje za ćelijskim zidom, što se naziva plazmoliza. Kada se ćelija stavi u rastvor manje koncentriran od citoplazme, dolazi do transporta vode u suprotnom smeru - u ćeliju. Međutim, postoje ograničenja za rastezljivost citoplazmatske membrane, a životinjska stanica na kraju pukne, dok biljna stanica to ne dozvoljava zbog svog snažnog ćelijskog zida. Fenomen punjenja čitavog unutrašnjeg prostora ćelije ćelijskim sadržajem naziva se deplazmoliza. Prilikom pripreme lijekova, posebno za intravensku primjenu, potrebno je uzeti u obzir intracelularnu koncentraciju soli, jer to može dovesti do oštećenja krvnih stanica (za to se koristi fiziološka otopina koncentracije 0,9% natrijevog klorida). To nije ništa manje važno kada se uzgajaju ćelije i tkiva, kao i životinjski i biljni organi.

Aktivan transport nastavlja sa trošenjem energije ATP-a od niže koncentracije supstance ka većoj. Izvodi se pomoću posebnih pumpnih proteina. Proteini pumpaju K+, Na+, Ca 2+ i druge jone kroz membranu, što pospješuje transport esencijalnih organskih tvari, kao i nastanak nervnih impulsa itd.
Endocitoza- ovo je aktivni proces apsorpcije tvari u ćeliji, u kojem membrana formira invaginacije, a zatim formira membranske vezikule - fagozomi, koji sadrže apsorbirane objekte. Tada se primarni lizozom spaja sa fagozomom i formira sekundarni lizozom, ili fagolizozom, ili digestivna vakuola. Sadržaj vezikule probavljaju enzimi lizosoma, a produkte razgradnje ćelija apsorbuje i asimilira. Nesvareni ostaci se uklanjaju iz ćelije egzocitozom. Postoje dvije glavne vrste endocitoze: fagocitoza i pinocitoza.
Fagocitoza je proces hvatanja na površini ćelije i apsorpcije čvrstih čestica od strane ćelije, i pinocitoza- tečnosti. Fagocitoza se javlja uglavnom u životinjskim ćelijama (jednoćelijske životinje, ljudski leukociti), obezbeđuje njihovu ishranu i često štiti organizam. Pinocitozom, proteini, kompleksi antigen-antitelo se apsorbuju tokom imunoloških reakcija, itd. Međutim, mnogi virusi ulaze u ćeliju i pinocitozom ili fagocitozom. U biljnim i gljivičnim stanicama fagocitoza je praktički nemoguća, jer su okružene trajnim ćelijskim membranama.

Egzocitoza- proces obrnut od endocitoze. Na taj način se iz probavnih vakuola oslobađaju nesvareni ostaci hrane, a uklanjaju se tvari neophodne za život stanice i tijela u cjelini. Na primjer, do prijenosa nervnih impulsa dolazi zbog oslobađanja hemijskih glasnika od strane neurona koji šalje impuls - posrednici, a u biljnim ćelijama na taj način se luče pomoćni ugljikohidrati stanične membrane.
Ćelijski zidovi biljnih ćelija, gljivica i bakterija. Izvan membrane, ćelija može lučiti jak okvir - stanične membrane, ili ćelijski zid.
Kod biljaka je osnova ćelijskog zida celuloza, upakovane u snopove od 50-100 molekula. Prostori između njih su ispunjeni vodom i drugim ugljikohidratima. Zid biljne ćelije prožet je tubulima - plazmodesmata, kroz koji prolaze membrane endoplazmatskog retikuluma. Plazmodezma obavlja transport tvari između stanica. Međutim, transport supstanci, kao što je voda, može se desiti i duž samih ćelijskih zidova. S vremenom se u ćelijskom zidu biljaka nakupljaju različite tvari, uključujući tanine ili tvari slične mastima, što dovodi do lignifikacije ili suberizacije samog ćelijskog zida, istiskivanja vode i odumiranja ćelijskog sadržaja. Između ćelijskih zidova susjednih biljnih ćelija nalaze se žele poput odstojnika - srednje ploče koje ih drže zajedno i cementiraju biljno tijelo kao cjelinu. Uništavaju se samo u procesu sazrevanja plodova i kada lišće opada.

Formiraju se ćelijski zidovi ćelija gljivica hitin- ugljeni hidrat koji sadrži azot. Prilično su jaki i vanjski su skelet ćelije, ali ipak, kao i kod biljaka, sprječavaju fagocitozu.
U bakterijama, stanični zid sadrži ugljikohidrate s peptidnim fragmentima - murein, međutim, njegov sadržaj značajno varira među različitim grupama bakterija. Drugi polisaharidi se također mogu lučiti na vrhu ćelijskog zida, formirajući mukoznu kapsulu koja štiti bakterije od vanjskih utjecaja.
Membrana određuje oblik ćelije, služi kao mehanički oslonac, obavlja zaštitnu funkciju, obezbeđuje osmotska svojstva ćelije, ograničavajući rastezanje živog sadržaja i sprečavajući rupturu ćelije koja se povećava usled ulaska vode. . Osim toga, voda i tvari otopljene u njoj prevladavaju ćelijski zid prije ulaska u citoplazmu ili, obrnuto, pri izlasku iz nje, dok se voda transportuje kroz ćelijske zidove brže nego kroz citoplazmu.
Citoplazma
Citoplazma- Ovo je unutrašnji sadržaj ćelije. Sve ćelijske organele, jezgro i razni otpadni proizvodi su uronjeni u njega.
Citoplazma povezuje sve dijelove ćelije jedni s drugima i u njoj se odvijaju brojne metaboličke reakcije. Citoplazma je odvojena od okoline i membranama podijeljena na odjeljke, odnosno ćelije imaju membransku strukturu. Može biti u dva stanja - sol i gel. Sol- ovo je polutečno, želeasto stanje citoplazme, u kojem se vitalni procesi odvijaju najintenzivnije, i gel- gušće, želatinasto stanje koje ometa nastanak hemijskih reakcija i transport supstanci.
Tečni dio citoplazme bez organela naziva se hijaloplazma. Hijaloplazma ili citosol je koloidna otopina u kojoj se nalazi neka vrsta suspenzije prilično velikih čestica, na primjer proteina, okruženih dipolima molekula vode. Taloženje ove suspenzije ne dolazi zbog činjenice da imaju isti naboj i da se međusobno odbijaju.
Organoidi
Organoidi- To su trajne komponente ćelije koje obavljaju određene funkcije.
Ovisno o strukturnim karakteristikama, dijele se na membranske i nemembranske. Membrane organele su zauzvrat klasifikovane kao jednomembranske (endoplazmatski retikulum, Golgijev kompleks i lizozomi) ili dvostruke membrane (mitohondrije, plastidi i jezgra). Nemembranski Organele su ribozomi, mikrotubule, mikrofilamenti i ćelijski centar. Od navedenih organela, samo ribozomi su inherentni prokariotima.
Struktura i funkcije jezgra. Core- velika dvomembranska organela koja leži u centru ćelije ili na njenoj periferiji. Dimenzije jezgra mogu biti u rasponu od 3-35 mikrona. Oblik jezgra je najčešće sferičan ili elipsoidan, ali postoje i štapićasta, vretenasta, zrnasta, režnjeva pa čak i segmentirana jezgra. Neki istraživači vjeruju da oblik jezgra odgovara obliku same ćelije.
Većina ćelija ima jedno jezgro, ali, na primer, u ćelijama jetre i srca mogu ih biti dva, au nizu neurona - do 15. Vlakna skeletnih mišića obično sadrže mnogo jezgara, ali to nisu ćelije u punom smislu te riječi, budući da nastaju kao rezultat fuzije nekoliko ćelija.
Jezgro je okruženo nuklearni omotač, a njen unutrašnji prostor je ispunjen nuklearni sok, ili nukleoplazma (karioplazma), u koji su uronjeni hromatin I nucleolus. Jezgro obavlja tako važne funkcije kao što su pohranjivanje i prijenos nasljednih informacija, kao i kontrola života ćelije.

Uloga jezgra u prijenosu nasljednih informacija uvjerljivo je dokazana u eksperimentima sa zelenom algom Acetabularia. U jednoj divovskoj ćeliji, koja doseže dužinu od 5 cm, razlikuju se kapa, stabljika i rizoid. Štaviše, sadrži samo jedno jezgro smješteno u rizoidu. Tridesetih godina I. Hemmerling je presadio jezgro jedne vrste acetabularia zelene boje u rizoid druge vrste, braon boje, iz koje je jezgro uklonjeno. Nakon nekog vremena, biljci sa presađenim jezgrom izrasla je nova kapica, poput alge donora jezgra. U isto vrijeme, klobuk ili stabljika, odvojena od rizoida i koja ne sadrži jezgro, nakon nekog vremena je umrla.

Nuklearni omotač formirane od dvije membrane - vanjske i unutrašnje, između kojih postoji prostor. Intermembranski prostor komunicira sa šupljinom grubog endoplazmatskog retikuluma, a vanjska membrana jezgra može nositi ribozome. Nuklearni omotač je prožet brojnim porama obloženim posebnim proteinima. Transport supstanci se odvija kroz pore: potrebni proteini (uključujući enzime), ioni, nukleotidi i druge supstance ulaze u jezgro, a molekule RNK, istrošeni proteini i podjedinice ribosoma ga napuštaju. Dakle, funkcije nuklearnog omotača su odvajanje sadržaja jezgre od citoplazme, kao i regulacija metabolizma između jezgre i citoplazme.
Nukleoplazma naziva se sadržaj jezgra, u koji su uronjeni hromatin i nukleolus. To je koloidna otopina, kemijski podsjeća na citoplazmu. Enzimi nukleoplazme kataliziraju razmjenu aminokiselina, nukleotida, proteina itd. Nukleoplazma je povezana s hijaloplazmom preko nuklearnih pora. Funkcije nukleoplazme, kao i hijaloplazme, su da osigura međusobnu povezanost svih strukturnih komponenti jezgra i da provede niz enzimskih reakcija.
Chromatin nazvana kolekcija tankih filamenata i granula uronjenih u nukleoplazmu. Može se otkriti samo bojenjem, jer su indeksi loma kromatina i nukleoplazme približno isti. Filamentna komponenta hromatina se naziva euhromatin, i granulirani - heterohromatin. Euhromatin je slabo zbijen, jer se iz njega čitaju nasljedne informacije, dok je spiraliziraniji heterohromatin genetski neaktivan.
Hromatin je strukturna modifikacija hromozoma u jezgri koja se ne dijeli. Dakle, kromosomi su stalno prisutni u jezgri, samo se njihovo stanje mijenja ovisno o funkciji koju jezgro u ovom trenutku obavlja.
Sastav hromatina uglavnom uključuje nukleoproteinske proteine (deoksiribonukleoproteine i ribonukleoproteine), kao i enzime, od kojih su najvažniji povezani sa sintezom nukleinskih kiselina, i nekih drugih supstanci.
Funkcije hromatina sastoje se, prvo, u sintezi nukleinskih kiselina specifičnih za dati organizam, koje usmjeravaju sintezu specifičnih proteina, i drugo, u prijenosu nasljednih svojstava sa ćelije majke na ćelije kćeri, za šta se hromatinske niti se pakuju u hromozome tokom procesa deobe.
Nucleolus- sferično tijelo, jasno vidljivo pod mikroskopom, prečnika 1-3 mikrona. Formira se na dijelovima kromatina u kojima su kodirane informacije o strukturi rRNA i ribosomskih proteina. U jezgru se često nalazi samo jedna jezgra, ali u onim stanicama u kojima se odvijaju intenzivni vitalni procesi mogu biti dvije ili više jezgara. Funkcije nukleola su sinteza rRNA i sastavljanje ribosomskih podjedinica kombinovanjem rRNK sa proteinima koji dolaze iz citoplazme.
Mitohondrije- dvomembranske organele okruglog, ovalnog ili štapićastog oblika, iako se nalaze i spiralne (u spermi). Prečnik mitohondrija je do 1 µm, a dužina do 7 µm. Prostor unutar mitohondrija ispunjen je matriksom. Matrix- Ovo je glavna supstanca mitohondrija. U nju su uronjeni kružni DNK molekul i ribozomi. Vanjska membrana mitohondrija je glatka i nepropusna za mnoge tvari. Unutrašnja membrana ima izbočine - cristas, povećavajući površinu membrane za hemijske reakcije. Na površini membrane nalaze se brojni proteinski kompleksi koji čine takozvani respiratorni lanac, kao i enzimi ATP sintetaze u obliku gljive. Aerobna faza disanja javlja se u mitohondrijima, tokom koje se sintetiše ATP.

Plastidi- velike dvomembranske organele, karakteristične samo za biljne ćelije. Unutrašnji prostor plastida je ispunjen stroma, ili matrica. Stroma sadrži manje ili više razvijen sistem membranskih vezikula - tilakoidi, koji se skupljaju u gomile - zrna, kao i vlastiti kružni DNK molekul i ribozomi. Postoje četiri glavne vrste plastida: hloroplasti, hromoplasti, leukoplasti i proplastidi.
Hloroplasti- to su zeleni plastidi promjera 3-10 mikrona, jasno vidljivi pod mikroskopom. Ima ih samo u zelenim dijelovima biljaka - listovima, mladim stabljikama, cvjetovima i plodovima. Kloroplasti su općenito ovalnog ili elipsoidnog oblika, ali također mogu biti u obliku čaše, spirale ili čak režnjevi. Broj hloroplasta u ćeliji je u prosjeku od 10 do 100 komada. Međutim, na primjer, u nekim algama može biti jedna, imati značajne dimenzije i složen oblik - tada se zove hromatofora. U drugim slučajevima, broj hloroplasta može doseći nekoliko stotina, dok su njihove veličine male. Boja hloroplasta je zbog glavnog pigmenta fotosinteze - hlorofil, iako sadrže i dodatne pigmente - karotenoidi. Karotenoidi postaju vidljivi tek u jesen, kada se hlorofil u ostarjelim listovima razgradi. Glavna funkcija hloroplasta je fotosinteza. Svjetlosne reakcije fotosinteze odvijaju se na tilakoidnim membranama na koje su pričvršćene molekule klorofila, a tamne reakcije se odvijaju u stromi, gdje se nalaze brojni enzimi.

Hromoplasti- To su žuti, narandžasti i crveni plastidi koji sadrže karotenoidne pigmente. Oblik hromoplasta također može značajno varirati: mogu biti cjevasti, sferni, kristalni, itd. Kromoplasti daju boju cvjetovima i plodovima biljaka, privlačeći oprašivače i distributere sjemena i plodova.
Leukoplasti- To su bijeli ili bezbojni plastidi, uglavnom okruglog ili ovalnog oblika. Uobičajeni su u nefotosintetičkim dijelovima biljaka, na primjer u kožici listova, gomoljima krompira itd. Pohranjuju hranjive tvari, najčešće škrob, ali kod nekih biljaka to mogu biti proteini ili ulje.
Plastidi se formiraju u biljnim ćelijama od proplastida, koji su već prisutni u ćelijama obrazovnog tkiva i predstavljaju mala dvomembranska tela. U ranim fazama razvoja, različite vrste plastida mogu se transformirati jedna u drugu: kada su izloženi svjetlosti, leukoplasti gomolja krumpira i hromoplasti korijena mrkve postaju zeleni.
Plastidi i mitohondrije nazivaju se poluautonomnim organelama ćelije, jer imaju sopstvene molekule DNK i ribozome, vrše sintezu proteina i dele se nezavisno od deobe ćelije. Ove karakteristike se objašnjavaju njihovim porijeklom od jednoćelijskih prokariotskih organizama. Međutim, „nezavisnost“ mitohondrija i plastida je ograničena, jer njihova DNK sadrži premalo gena za slobodno postojanje, dok je ostatak informacija kodiran u hromozomima jezgre, što mu omogućava da kontroliše ove organele.
Endoplazmatski retikulum (ER), ili endoplazmatski retikulum (ER), je jednomembranska organela, koja je mreža membranskih šupljina i tubula koji zauzimaju do 30% sadržaja citoplazme. Prečnik EPS tubula je oko 25-30 nm. Postoje dve vrste EPS-a - grubi i glatki. Rough XPS nosi ribozome, gdje se odvija sinteza proteina. Smooth XPS nema ribozoma. Njegova funkcija je sinteza lipida i ugljikohidrata, kao i transport, skladištenje i neutralizacija toksičnih tvari. Posebno je razvijen u onim stanicama u kojima se odvijaju intenzivni metabolički procesi, na primjer u ćelijama jetre - hepatocitima - i skeletnim mišićnim vlaknima. Supstance sintetizovane u ER transportuju se do Golgijevog aparata. Sastavljanje ćelijskih membrana se također događa u ER-u, ali se njihovo formiranje završava u Golgijevom aparatu.
Golgijev aparat, ili Golgijev kompleks, je jednomembranska organela formirana sistemom ravnih cisterni, tubula i vezikula odvojenih od njih. Strukturna jedinica Golgijevog aparata je dictyosome- gomila rezervoara, na čiji jedan pol dolaze supstance iz EPS-a, a sa suprotnog pola, podvrgnuti određenim transformacijama, pakuju se u vezikule i šalju u druge delove ćelije. Prečnik rezervoara je oko 2 mikrona, a prečnik malih mehurića je oko 20-30 mikrona. Glavne funkcije Golgijevog kompleksa su sinteza određenih supstanci i modifikacija (promjena) proteina, lipida i ugljikohidrata koji dolaze iz ER, konačno formiranje membrana, kao i transport tvari kroz ćeliju, obnavljanje njenih struktura. i formiranje lizosoma. Golgijev aparat dobio je ime po italijanskom naučniku Camillu Golgiju, koji je prvi otkrio ovu organelu (1898).

Lizozomi- male jednomembranske organele do 1 μm u promjeru, koje sadrže hidrolitičke enzime uključene u unutarćelijsku probavu. Membrane lizosoma su slabo propusne za ove enzime, pa lizozomi vrlo precizno i ciljano obavljaju svoje funkcije. Tako aktivno učestvuju u procesu fagocitoze, formirajući probavne vakuole, a u slučaju izgladnjivanja ili oštećenja pojedinih dijelova stanice, probavljaju ih bez utjecaja na druge. Nedavno je otkrivena uloga lizosoma u procesima stanične smrti.
Vacuole je šupljina u citoplazmi biljnih i životinjskih ćelija, omeđena membranom i ispunjena tečnošću. Digestivne i kontraktilne vakuole nalaze se u ćelijama protozoa. Prvi učestvuju u procesu fagocitoze, jer razgrađuju hranljive materije. Potonji osiguravaju održavanje ravnoteže vode i soli zbog osmoregulacije. Kod višećelijskih životinja uglavnom se nalaze probavne vakuole.
U biljnim ćelijama vakuole su uvijek prisutne, okružene su posebnom membranom i ispunjene ćelijskim sokom. Membrana koja okružuje vakuolu slična je po hemijskom sastavu, strukturi i funkcijama plazma membrani. Ćelijski sok je vodeni rastvor različitih neorganskih i organskih supstanci, uključujući mineralne soli, organske kiseline, ugljene hidrate, proteine, glikozide, alkaloide itd. Vakuola može zauzeti do 90% zapremine ćelije i potisnuti jezgro ka periferiji. Ovaj dio ćelije vrši skladištenje, izlučivanje, osmotsku, zaštitnu, lizozomsku i druge funkcije, jer akumulira hranjive tvari i otpadne tvari, osigurava snabdijevanje vodom i održava oblik i volumen ćelije, a sadrži i enzime za razgradnju mnoge ćelijske komponente. Osim toga, biološki aktivne tvari vakuola mogu spriječiti mnoge životinje da jedu ove biljke. Kod brojnih biljaka zbog bubrenja vakuola dolazi do rasta ćelija produljenjem.
Vakuole su prisutne i u ćelijama nekih gljiva i bakterija, ali kod gljiva obavljaju samo funkciju osmoregulacije, dok kod cijanobakterija održavaju plovnost i učestvuju u procesu asimilacije dušika iz zraka.
Ribosomi- male nemembranske organele promjera 15-20 mikrona, koje se sastoje od dvije podjedinice - velike i male. Eukariotske ribosomske podjedinice se sklapaju u nukleolu i zatim transportuju u citoplazmu. Ribosomi kod prokariota, mitohondrija i plastida su manje veličine od ribozoma u eukariota. Ribosomalne podjedinice uključuju rRNA i proteine.

Broj ribozoma u ćeliji može doseći nekoliko desetina miliona: u citoplazmi, mitohondrijima i plastidima oni su u slobodnom stanju, a na grubom ER - u vezanom stanju. Oni učestvuju u sintezi proteina, posebno provode proces translacije - biosintezu polipeptidnog lanca na molekulu mRNA. Slobodni ribozomi sintetiziraju proteine hijaloplazme, mitohondrija, plastida i vlastite ribosomske proteine, dok ribozomi vezani za grubi ER vrše translaciju proteina za uklanjanje iz ćelija, sklapanje membrane i formiranje lizosoma i vakuola.
Ribosomi se mogu naći pojedinačno u hijaloplazmi ili sastavljeni u grupe tokom simultane sinteze nekoliko polipeptidnih lanaca na jednoj mRNK. Takve grupe ribozoma se nazivaju poliribozomi, ili polizomi.

Mikrotubule- To su cilindrične šuplje nemembranske organele koje prodiru kroz cijelu citoplazmu ćelije. Njihov promjer je oko 25 nm, debljina stijenke 6-8 nm. Formirani su od brojnih proteinskih molekula tubulin, koje prvo formiraju 13 niti koje liče na perle, a zatim se sklapaju u mikrotubulu. Mikrotubule formiraju citoplazmatski retikulum, koji ćeliji daje oblik i zapreminu, povezuje plazma membranu sa ostalim delovima ćelije, obezbeđuje transport supstanci kroz ćeliju, učestvuje u kretanju ćelije i intracelularnih komponenti, kao i u podjela genetskog materijala. Oni su dio ćelijskog centra i organela kretanja - flagela i cilija.
mikrofilamenti, ili microthreads, su također nemembranske organele, međutim, imaju filamentasti oblik i formiraju ih ne tubulin, već actin. Učestvuju u procesima membranskog transporta, intercelularnog prepoznavanja, diobe ćelijske citoplazme i njenog kretanja. U mišićnim ćelijama, interakcija aktinskih mikrofilamenata sa filamentima miozina posreduje u kontrakciji.
Mikrotubule i mikrofilamenti čine unutrašnji skelet ćelije - citoskelet. To je složena mreža vlakana koja pružaju mehaničku potporu plazma membrani, određuju oblik ćelije, lokaciju ćelijskih organela i njihovo kretanje tokom ćelijske deobe.

Ćelijski centar- nemembranska organela koja se nalazi u životinjskim ćelijama u blizini jezgra; nema ga u biljnim ćelijama. Dužina mu je oko 0,2-0,3 mikrona, a prečnik 0,1-0,15 mikrona. Ćelijski centar formiraju dva centriola, koji leže u međusobno okomitim ravninama, i radijantna sfera iz mikrotubula. Svaki centriol je formiran od devet grupa mikrotubula, sakupljenih u grupe od tri, odnosno tripleta. Ćelijski centar učestvuje u procesima sklapanja mikrotubula, deobi naslednog materijala ćelije, kao i u formiranju bičaka i cilija.

Organele kretanja. Flagella I cilia Oni su izrasline ćelije prekrivene plazmalemom. Osnovu ovih organela čini devet pari mikrotubula smještenih duž periferije i dvije slobodne mikrotubula u centru. Mikrotubule su međusobno povezane različitim proteinima, osiguravajući njihovo koordinirano odstupanje od ose - oscilacije. Oscilacije su energetski zavisne, odnosno na ovaj proces se troši energija visokoenergetskih ATP veza. Obnavljanje izgubljenih flagela i cilija je funkcija bazalna tijela, ili kinetozomi nalazi u njihovoj bazi.
Dužina cilija je oko 10-15 nm, a dužina flagela je 20-50 µm. Zbog striktno usmjerenih pokreta bičaka i cilija ne dolazi samo do kretanja jednoćelijskih životinja, sperme i sl., već i do čišćenja disajnih puteva i kretanja jajne stanice kroz jajovode, budući da svi ovi dijelovi ljudskog tijela obložene su trepljastim epitelom.

Inkluzije
Inkluzije- To su nestalne komponente ćelije koje se formiraju i nestaju tokom njenog života. To uključuje i rezervne tvari, na primjer, zrnca škroba ili proteina u biljnim stanicama, granule glikogena u stanicama životinja i gljiva, volutin u bakterijama, kapi masti u svim vrstama stanica, i otpadne proizvode, posebno ostatke hrane. nesvareni kao rezultat fagocitoze, formirajući takozvana rezidualna tijela.

Odnos između strukture i funkcija dijelova i organela ćelije je osnova njenog integriteta
Svaki od dijelova ćelije, s jedne strane, je posebna struktura sa specifičnom strukturom i funkcijama, as druge, komponenta složenijeg sistema koji se naziva ćelija. Većina nasljednih informacija eukariotske stanice koncentrirana je u jezgru, ali samo jezgro nije u stanju osigurati njegovu implementaciju, jer je za to potrebna barem citoplazma, koja djeluje kao glavna tvar, i ribozomi na kojima se odvija ova sinteza. . Većina ribozoma se nalazi na granularnom endoplazmatskom retikulumu, odakle se proteini najčešće transportuju do Golgijevog kompleksa, a zatim, nakon modifikacije, do onih dijelova ćelije za koje su namijenjeni, odnosno izlučuju se. Membransko pakovanje proteina i ugljikohidrata može se ugraditi u membrane organela i citoplazmatsku membranu, osiguravajući njihovo stalno obnavljanje. Lizozomi i vakuole, koji obavljaju važne funkcije, također se odvajaju od Golgijevog kompleksa. Na primjer, bez lizosoma, stanice bi se brzo pretvorile u svojevrsno deponije za otpadne molekule i strukture.
Za nastanak svih ovih procesa potrebna je energija koju proizvode mitohondriji, au biljkama hloroplasti. I iako su ove organele relativno autonomne, budući da imaju svoje molekule DNK, neki od njihovih proteina su još uvijek kodirani nuklearnim genomom i sintetizirani u citoplazmi.
Dakle, ćelija je neraskidivo jedinstvo svojih sastavnih komponenti, od kojih svaka obavlja svoju jedinstvenu funkciju.
Metabolizam i pretvaranje energije svojstva su živih organizama. Energetski i plastični metabolizam, njihov odnos. Faze energetskog metabolizma. Fermentacija i disanje. Fotosinteza, njen značaj, kosmička uloga. Faze fotosinteze. Reakcije svjetlosti i tame fotosinteze, njihov odnos. Hemosinteza. Uloga hemosintetskih bakterija na Zemlji
Metabolizam i pretvaranje energije - svojstva živih organizama
Ćelija se može uporediti sa minijaturnom hemijskom fabrikom u kojoj se dešavaju stotine i hiljade hemijskih reakcija.
Metabolizam- skup hemijskih transformacija u cilju očuvanja i samoreprodukcije bioloških sistema.
Uključuje unos supstanci u organizam tokom ishrane i disanja, unutarćelijskog metabolizma ili metabolizam, kao i izolaciju konačnih metaboličkih proizvoda.
Metabolizam je neraskidivo povezan sa procesima pretvaranja jedne vrste energije u drugu. Na primjer, tokom procesa fotosinteze svjetlosna energija se pohranjuje u obliku energije hemijskih veza složenih organskih molekula, a tokom procesa disanja se oslobađa i troši na sintezu novih molekula, mehanički i osmotski rad, raspršen u obliku toplote itd.
Pojava hemijskih reakcija u živim organizmima osigurana je zahvaljujući biološkim katalizatorima proteinske prirode - enzimi, ili enzimi. Kao i drugi katalizatori, enzimi ubrzavaju nastanak hemijskih reakcija u ćeliji za desetine i stotine hiljada puta, a ponekad ih čak i omogućavaju, ali ne menjaju prirodu ili svojstva krajnjeg(ih) proizvoda(a) reakcije i čine ne menjaju sebe. Enzimi mogu biti i jednostavni i složeni proteini, koji osim proteinskog, uključuju i neproteinski dio - kofaktor (koenzim). Primjeri enzima su pljuvačka amilaza, koja razgrađuje polisaharide tokom dužeg žvakanja, i pepsin, koji osigurava varenje proteina u želucu.
Enzimi se razlikuju od neproteinskih katalizatora po visokoj specifičnosti djelovanja, značajnom povećanju brzine reakcije uz njihovu pomoć, kao i po sposobnosti reguliranja djelovanja promjenom uvjeta reakcije ili interakcije različitih tvari s njima. Osim toga, uvjeti pod kojima se odvija enzimska kataliza značajno se razlikuju od onih pod kojima se odvija neenzimska kataliza: optimalna temperatura za funkcioniranje enzima u ljudskom tijelu je $37°C$, tlak treba biti blizu atmosferskog, a $pH$ okoline može značajno oklevati. Dakle, amilaza zahtijeva alkalno okruženje, a pepsin kiselo okruženje.
Mehanizam djelovanja enzima je smanjenje energije aktivacije supstanci (supstrata) koje ulaze u reakciju zbog stvaranja intermedijarnih kompleksa enzim-supstrat.

Energetski i plastični metabolizam, njihov odnos
Metabolizam se sastoji od dva procesa koja se odvijaju istovremeno u ćeliji: plastičnog i energetskog metabolizma.
Plastični metabolizam (anabolizam, asimilacija) je skup reakcija sinteze koje uključuju trošenje ATP energije. U procesu plastičnog metabolizma sintetiziraju se organske tvari potrebne za ćeliju. Primjeri reakcija plastične razmjene su fotosinteza, biosinteza proteina i replikacija DNK (samo-duplikacija).
Energetski metabolizam (katabolizam, disimilacija) je skup reakcija koje razlažu složene supstance na jednostavnije. Kao rezultat energetskog metabolizma, energija se oslobađa i pohranjuje u obliku ATP-a. Najvažniji procesi energetskog metabolizma su disanje i fermentacija.
Plastična i energetska razmjena su neraskidivo povezane, jer se u procesu plastične izmjene sintetiziraju organske tvari i za to je potrebna energija ATP-a, a u procesu izmjene energije organske tvari se razgrađuju i oslobađa energija koja će se potom trošiti na procese sinteze. .
Organizmi primaju energiju tokom procesa ishrane, a oslobađaju je i pretvaraju u pristupačan oblik uglavnom tokom procesa disanja. Prema načinu ishrane svi organizmi se dele na autotrofe i heterotrofe. Autotrofi sposoban da samostalno sintetiše organske supstance od neorganskih, i heterotrofi koristiti isključivo pripremljene organske supstance.
Faze energetskog metabolizma
Unatoč složenosti reakcija energetskog metabolizma, on se konvencionalno dijeli u tri faze: pripremni, anaerobni (bez kisika) i aerobni (kiseonik).
On pripremna faza molekule polisaharida, lipida, proteina, nukleinskih kiselina razlažu se na jednostavnije, na primjer glukozu, glicerol i masne kiseline, aminokiseline, nukleotide itd. Ova faza se može javiti direktno u stanicama ili u crijevima, odakle dolazi do razgradnje. donje supstance se isporučuju kroz krvotok.
Anaerobna faza energetski metabolizam je praćen daljnjim razlaganjem monomera organskih spojeva u još jednostavnije međuproizvode, na primjer, pirogrožđanu kiselinu ili piruvat. Ne zahtijeva prisustvo kisika, a za mnoge organizme koji žive u blatu močvara ili u ljudskim crijevima, to je jedini način da dobiju energiju. Anaerobna faza energetskog metabolizma odvija se u citoplazmi.
Različite tvari mogu biti podvrgnute cijepanju bez kisika, ali vrlo često je supstrat reakcija glukoza. Proces njegovog cijepanja bez kisika naziva se glikoliza. Tokom glikolize, molekul glukoze gubi četiri atoma vodika, odnosno oksidira se i nastaju dva molekula pirogrožđane kiseline, dva molekula ATP-a i dva molekula redukovanog nosača vodonika $NADH + H^(+)$:
$C_6H_(12)O_6 + 2H_3PO_4 + 2ADP + 2NAD → 2C_3H_4O_3 + 2ATP + 2NADH + H^(+) + 2H_2O$.
Do stvaranja ATP-a iz ADP-a dolazi zbog direktnog prijenosa fosfatnog anjona iz predfosforiliranog šećera i naziva se fosforilacija supstrata.
Aerobna faza do razmjene energije može doći samo u prisustvu kisika, dok se međuspojni spojevi koji nastaju cijepanjem bez kisika oksidiraju do konačnih proizvoda (ugljični dioksid i voda) i oslobađa se najveći dio energije pohranjene u kemijskim vezama organskih spojeva. Pretvara se u energiju visokoenergetskih veza od 36 ATP molekula. Ova faza se još naziva tkivno disanje. U nedostatku kisika, intermedijarni spojevi se pretvaraju u druge organske tvari, proces tzv fermentacija.
Dah
Mehanizam ćelijskog disanja šematski je prikazan na Sl.

Aerobno disanje se događa u mitohondrijama, pri čemu pirogrožđana kiselina prvo gubi jedan atom ugljika, što je praćeno sintezom jednog redukcijskog ekvivalenta $NADH + H^(+)$ i molekula acetil koenzima A (acetil-CoA):
$C_3H_4O_3 + NAD + H~CoA → CH_3CO~CoA + NADH + H^(+) + CO_2$.
Acetil-CoA u mitohondrijskom matriksu je uključen u lanac hemijskih reakcija, čija se ukupnost naziva Krebsov ciklus (ciklus trikarboksilne kiseline, ciklus limunske kiseline). Tokom ovih transformacija formiraju se dva ATP molekula, acetil-CoA se potpuno oksidira u ugljični dioksid, a njegovi vodikovi joni i elektroni se dodaju na nosače vodonika $NADH + H^(+)$ i $FADH_2$. Nosači prenose vodikove protone i elektrone do unutrašnjih membrana mitohondrija, formirajući kriste. Uz pomoć proteina nosača, protoni vodonika se pumpaju u međumembranski prostor, a elektroni se prenose kroz tzv. respiratorni lanac enzima koji se nalaze na unutrašnjoj membrani mitohondrija i ispuštaju se na atome kiseonika:
$O_2+2e^(-)→O_2^-$.
Treba napomenuti da neki proteini respiratornog lanca sadrže željezo i sumpor.
Iz intermembranskog prostora protoni vodika se transportuju nazad u mitohondrijsku matricu uz pomoć posebnih enzima - ATP sintaza, a energija koja se pri tome oslobađa troši se na sintezu 34 ATP molekula iz svakog molekula glukoze. Ovaj proces se zove oksidativna fosforilacija. U mitohondrijskom matriksu, vodikovi protoni reaguju sa radikalima kiseonika i formiraju vodu:
$4H^(+)+O_2^-→2H_2O$.
Skup reakcija disanja kisika može se izraziti na sljedeći način:
$2C_3H_4O_3 + 6O_2 + 36H_3PO_4 + 36ADP → 6CO_2 + 38H_2O + 36ATP.$
Ukupna jednačina disanja izgleda ovako:
$C_6H_(12)O_6 + 6O_2 + 38H_3PO_4 + 38ADP → 6CO_2 + 40H_2O + 38ATP.$
Fermentacija
U nedostatku kiseonika ili njegovom nedostatku dolazi do fermentacije. Fermentacija je evolucijski ranija metoda dobivanja energije od disanja, ali je energetski manje korisna jer fermentacija proizvodi organske tvari koje su još uvijek bogate energijom. Postoji nekoliko glavnih tipova fermentacije: mliječna, alkoholna, octena itd. Tako se u skeletnim mišićima u nedostatku kisika tokom fermentacije, pirogrožđana kiselina reducira u mliječnu kiselinu, dok se prethodno formirani redukcijski ekvivalenti troše, a samo ostaju dva ATP molekula:
$2C_3H_4O_3 + 2NADH + H^(+) → 2C_3H_6O_3 + 2NAD$.
Tokom fermentacije uz pomoć kvasca, pirogrožđana kiselina se u prisustvu kiseonika pretvara u etil alkohol i ugljen monoksid (IV):
$C_3H_4O_3 + NADH + H^(+) → C_2H_5OH + CO_2 + NAD^(+)$.
Prilikom fermentacije uz pomoć mikroorganizama iz pirogrožđane kiseline mogu nastati i sirćetna, maslačna, mravlja kiselina itd.
ATP, dobijen kao rezultat energetskog metabolizma, troši se u ćeliji za različite vrste rada: hemijski, osmotski, električni, mehanički i regulatorni. Hemijski rad uključuje biosintezu proteina, lipida, ugljikohidrata, nukleinskih kiselina i drugih vitalnih spojeva. Osmotski rad uključuje procese apsorpcije ćelije i uklanjanja iz nje supstanci koje se nalaze u ekstracelularnom prostoru u koncentracijama većim nego u samoj ćeliji. Električni rad je usko povezan s osmotskim radom, jer se kao rezultat kretanja nabijenih čestica kroz membrane formira naboj membrane i stječu svojstva ekscitabilnosti i provodljivosti. Mehanički rad uključuje kretanje tvari i struktura unutar ćelije, kao i ćelije u cjelini. Regulatorni rad obuhvata sve procese koji imaju za cilj koordinaciju procesa u ćeliji.
Fotosinteza, njen značaj, kosmička uloga
fotosinteza je proces pretvaranja svjetlosne energije u energiju kemijskih veza organskih spojeva uz sudjelovanje hlorofila.
Kao rezultat fotosinteze, godišnje se proizvede oko 150 milijardi tona organske materije i oko 200 milijardi tona kiseonika. Ovaj proces osigurava kruženje ugljika u biosferi, sprječavajući nakupljanje ugljičnog dioksida i na taj način sprječavajući efekat staklene bašte i pregrijavanje Zemlje. Organske tvari nastale kao rezultat fotosinteze ne troše u potpunosti drugi organizmi, značajan dio njih je tokom miliona godina formirao naslage minerala (kameni i mrki ugalj, nafta). Nedavno su kao gorivo počeli da se koriste i repičino ulje („biodizel“) i alkohol dobijen iz biljnih ostataka. Ozon nastaje iz kiseonika pod uticajem električnih pražnjenja, koji formira ozonski ekran koji štiti sav život na Zemlji od razornog dejstva ultraljubičastih zraka.
Naš sunarodnik, izvanredni biljni fiziolog K. A. Timiryazev (1843-1920), nazvao je ulogu fotosinteze "kosmičkom", jer povezuje Zemlju sa Suncem (svemirom), osiguravajući priliv energije planeti.
Faze fotosinteze. Reakcije svjetlosti i tame fotosinteze, njihov odnos
Godine 1905. engleski biljni fiziolog F. Blackman otkrio je da se stopa fotosinteze ne može povećavati beskonačno; neki faktor je ograničava. Na osnovu toga, on je pretpostavio da postoje dvije faze fotosinteze: svjetlo I mračno. Pri niskom intenzitetu svjetlosti, brzina svjetlosnih reakcija raste proporcionalno porastu intenziteta svjetlosti, a osim toga, ove reakcije ne zavise od temperature, jer ne zahtijevaju enzime. Svjetlosne reakcije se javljaju na tilakoidnim membranama.
Brzina tamnih reakcija, naprotiv, raste sa povećanjem temperature, međutim, kada se postigne temperaturni prag od $30°C$, ovo povećanje prestaje, što ukazuje na enzimsku prirodu ovih transformacija koje se dešavaju u stromi. Treba napomenuti da svjetlost ima određeni utjecaj i na tamne reakcije, uprkos činjenici da se one nazivaju tamnim reakcijama.

Svjetlosna faza fotosinteze odvija se na tilakoidnim membranama koje nose nekoliko tipova proteinskih kompleksa, od kojih su glavni fotosistemi I i II, kao i ATP sintaza. Fotosistemi uključuju pigmentne komplekse, koji pored hlorofila sadrže i karotenoide. Karotenoidi hvataju svjetlost u područjima spektra gdje hlorofil nema, a također štite hlorofil od uništenja svjetlošću visokog intenziteta.
Pored pigmentnih kompleksa, fotosistemi uključuju i niz proteina akceptora elektrona, koji uzastopno prenose elektrone s molekula hlorofila jedan na drugi. Redoslijed ovih proteina se naziva lanac transporta elektrona hloroplasta.
Poseban kompleks proteina je takođe povezan sa fotosistemom II, koji obezbeđuje oslobađanje kiseonika tokom fotosinteze. Ovaj kompleks koji oslobađa kiseonik sadrži ione mangana i hlora.
IN svetlosna faza svjetlosni kvanti, odnosno fotoni, koji padaju na molekule klorofila smještene na tilakoidnim membranama, prenose ih u pobuđeno stanje, koje karakterizira veća energija elektrona. U ovom slučaju, pobuđeni elektroni iz hlorofila fotosistema I prenose se kroz lanac posrednika do nosača vodonika NADP, koji vezuje protone vodonika, uvek prisutne u vodenom rastvoru:
$NADP + 2e^(-) + 2H^(+) → NADPH + H^(+)$.
Redukovani $NADPH + H^(+)$ će se kasnije koristiti u mračnoj fazi. Elektroni iz hlorofila fotosistema II se takođe prenose duž lanca transporta elektrona, ali oni popunjavaju “elektronske rupe” hlorofila fotosistema I. Nedostatak elektrona u hlorofilu fotosistema II popunjava se oduzimanjem molekula vode, koje se dešava uz učešće već pomenutog kompleksa koji oslobađa kiseonik. Kao rezultat raspadanja molekula vode, što se tzv fotoliza, formiraju se protoni vodika i oslobađa se molekularni kiseonik, koji je nusproizvod fotosinteze:
$H_2O → 2H^(+) + 2e^(-) + (1)/(2)O_2$.
Genetske informacije u ćeliji. Geni, genetski kod i njegova svojstva. Matrična priroda reakcija biosinteze. Biosinteza proteina i nukleinskih kiselina
Genetske informacije u ćeliji
Reprodukcija svoje vrste jedno je od osnovnih svojstava živih bića. Zahvaljujući ovom fenomenu, postoji sličnost ne samo između organizama, već i između pojedinačnih ćelija, kao i njihovih organela (mitohondrija i plastida). Materijalna osnova ove sličnosti je prijenos genetskih informacija šifriranih u nukleotidnoj sekvenci DNK, koji se odvija kroz procese replikacije DNK (samo-duplikacije). Sve karakteristike i svojstva ćelija i organizama ostvaruju se zahvaljujući proteinima, čija je struktura prvenstveno određena slijedom nukleotida DNK. Stoga, biosinteza nukleinskih kiselina i proteina igra veliku važnost u metaboličkim procesima. Strukturna jedinica nasljedne informacije je gen.
Geni, genetski kod i njegova svojstva
Nasljedne informacije u ćeliji nisu monolitne, podijeljene su na zasebne "riječi" - gene.
Gene je elementarna jedinica genetske informacije.
Rad na programu “Ljudski genom”, koji je istovremeno sproveden u nekoliko zemalja i završen početkom ovog veka, dao nam je razumevanje da osoba ima samo oko 25-30 hiljada gena, ali informacije iz većine našeg DNK se nikada ne čita, jer sadrži ogroman broj besmislenih sekcija, ponavljanja i gena koji kodiraju osobine koje su izgubile značenje za ljude (rep, dlake na tijelu, itd.). Osim toga, dešifrovan je niz gena odgovornih za razvoj nasljednih bolesti, kao i ciljnih gena lijekova. Međutim, praktična primjena rezultata dobijenih tokom implementacije ovog programa odgađa se dok se ne dešifruju genomi većeg broja ljudi i ne postane jasno po čemu se razlikuju.
Geni koji kodiraju primarnu strukturu proteina, ribosomske ili transferne RNK se nazivaju strukturalni i geni koji omogućavaju aktivaciju ili supresiju čitanja informacija iz strukturnih gena - regulatorni. Međutim, čak i strukturni geni sadrže regulatorne regije.
Nasljedne informacije organizama su šifrirane u DNK u obliku određenih kombinacija nukleotida i njihovog niza - genetski kod. Njegova svojstva su: trostrukost, specifičnost, univerzalnost, redundantnost i nepreklapanje. Osim toga, u genetskom kodu nema znakova interpunkcije.
Svaka aminokiselina je kodirana u DNK sa tri nukleotida - trojka, na primjer, metionin je kodiran TAC tripletom, odnosno kod je triplet. S druge strane, svaki triplet kodira samo jednu aminokiselinu, što je njena specifičnost ili nedvosmislenost. Genetski kod je univerzalan za sve žive organizme, odnosno nasljedne informacije o ljudskim proteinima mogu čitati bakterije i obrnuto. Ovo ukazuje na jedinstvo porijekla organskog svijeta. Međutim, 64 kombinacije od tri nukleotida odgovaraju samo 20 aminokiselina, zbog čega jedna aminokiselina može biti kodirana sa 2-6 tripleta, odnosno genetski kod je suvišan ili degenerisan. Tri trojke nemaju odgovarajuće aminokiseline, nazivaju se stop kodoni, jer ukazuju na kraj sinteze polipeptidnog lanca.
Slijed baza u DNK tripletima i aminokiseline koje kodiraju

*Stop kodon, koji označava kraj sinteze polipeptidnog lanca.
Skraćenice za nazive aminokiselina:
Ala - alanin
Arg - arginin
Asn - asparagin
Asp - asparaginska kiselina
Val - valin
Njegov - histidin
Gly - glicin
Gln - glutamin
Glu - glutaminska kiselina
Ile - izoleucin
Leu - leucin
Liz - lizin
Met - metionin
Pro - proline
Ser - serin
Tyr - tirozin
Tre - treonin
Tri - triptofan
Fen - fenilalanin
Cys - cistein
Ako počnete čitati genetske informacije ne od prvog nukleotida u tripletu, već od drugog, onda će se okvir čitanja pomaknuti, već će i protein sintetiziran na ovaj način biti potpuno drugačiji ne samo u nukleotidnoj sekvenci, već iu struktura i svojstva. Između trojki nema znakova interpunkcije, tako da nema prepreka za pomicanje okvira čitanja, što otvara prostor za nastanak i održavanje mutacija.
Matrična priroda reakcija biosinteze
Bakterijske ćelije su sposobne da se udvostruče svakih 20-30 minuta, a eukariotske ćelije - svaki dan i još češće, što zahteva veliku brzinu i tačnost replikacije DNK. Osim toga, svaka stanica sadrži stotine i tisuće kopija mnogih proteina, posebno enzima, stoga je "parealni" način njihove proizvodnje neprihvatljiv za njihovu reprodukciju. Progresivnija metoda je žigosanje, koje vam omogućava da dobijete brojne točne kopije proizvoda i smanjite njegovu cijenu. Za štancanje je potrebna matrica iz koje se pravi otisak.
U ćelijama princip sinteze šablona je da se novi molekuli proteina i nukleinskih kiselina sintetišu u skladu sa programom ugrađenim u strukturu već postojećih molekula istih nukleinskih kiselina (DNK ili RNK).
Biosinteza proteina i nukleinskih kiselina
DNK replikacija. DNK je dvolančani biopolimer, čiji su monomeri nukleotidi. Kada bi se biosinteza DNK odvijala na principu fotokopiranja, neminovno bi se pojavila brojna izobličenja i greške u nasljednim informacijama, što bi u konačnici dovelo do smrti novih organizama. Stoga se proces udvostručavanja DNK odvija drugačije, na polukonzervativan način: molekul DNK se odmotava i na svakom od lanaca se sintetiše novi lanac prema principu komplementarnosti. Proces samoreprodukcije molekule DNK, koji osigurava precizno kopiranje nasljednih informacija i njihov prijenos s generacije na generaciju, naziva se replikacija(od lat. replicationo- ponavljanje). Kao rezultat replikacije, formiraju se dvije apsolutno točne kopije molekula DNK majke, od kojih svaka nosi po jednu kopiju molekula DNK majke.

Proces replikacije je zapravo izuzetno složen, budući da je u njemu uključen veliki broj proteina. Neki od njih odmotavaju dvostruku spiralu DNK, drugi razbijaju vodikove veze između nukleotida komplementarnih lanaca, treći (na primjer, enzim DNK polimeraza) odabiru nove nukleotide na principu komplementarnosti, itd. Dvije molekule DNK nastale kao rezultat replikacije se razilaze na dva tokom diobe novoformirane kćerke ćelije.
Greške u procesu replikacije nastaju izuzetno rijetko, ali ako se i dogode, vrlo brzo se eliminiraju kako DNK polimerazama tako i posebnim enzimima za popravak, jer svaka greška u nukleotidnoj sekvenci može dovesti do nepovratne promjene strukture i funkcija proteina. i, na kraju, negativno utiču na održivost nove ćelije ili čak pojedinca.
Biosinteza proteina. Kao što je figurativno rekao istaknuti filozof 19. veka F. Engels: „Život je oblik postojanja proteinskih tela“. Struktura i svojstva proteinskih molekula određuju se njihovom primarnom strukturom, odnosno sekvencom aminokiselina kodiranih u DNK. Od tačnosti reprodukcije ovih informacija zavisi ne samo postojanje samog polipeptida, već i funkcionisanje ćelije u celini, pa je proces sinteze proteina od velike važnosti. Čini se da je to najsloženiji proces sinteze u ćeliji, jer uključuje do tri stotine različitih enzima i drugih makromolekula. Osim toga, teče velikom brzinom, što zahtijeva još veću preciznost.
Postoje dvije glavne faze u biosintezi proteina: transkripcija i translacija.
Transkripcija(od lat. transkripcija- prepisivanje) je biosinteza mRNA molekula na DNK matrici.

Budući da molekula DNK sadrži dva antiparalelna lanca, čitanje informacija iz oba lanca dovelo bi do stvaranja potpuno različitih mRNA, pa je njihova biosinteza moguća samo na jednom od lanaca, koji se naziva kodirajući, odnosno kodogen, za razliku od drugog, nekodirajući ili nekodogeni. Proces ponovnog pisanja osigurava poseban enzim, RNA polimeraza, koji bira RNA nukleotide prema principu komplementarnosti. Ovaj proces se može dogoditi i u jezgri i u organelama koje imaju svoju DNK - mitohondrije i plastide.
Molekuli mRNA sintetizirani tokom transkripcije prolaze kroz složen proces pripreme za translaciju (mitohondrijalne i plastidne mRNA mogu ostati unutar organela, gdje se odvija druga faza biosinteze proteina). Tokom procesa sazrijevanja mRNA, za nju su vezana prva tri nukleotida (AUG) i rep adenil nukleotida, čija dužina određuje koliko kopija proteina može biti sintetizirano na datom molekulu. Tek tada zrele mRNA napuštaju jezgro kroz nuklearne pore.
Paralelno, u citoplazmi se odvija proces aktivacije aminokiselina, tokom kojeg se aminokiselina pridružuje odgovarajućoj slobodnoj tRNK. Ovaj proces katalizira poseban enzim i zahtijeva ATP.
Broadcast(od lat. emitovanje- transfer) je biosinteza polipeptidnog lanca na matrici mRNA, tokom koje se genetska informacija prevodi u aminokiselinsku sekvencu polipeptidnog lanca.

Druga faza sinteze proteina najčešće se javlja u citoplazmi, na primjer na grubom ER. Za njen nastanak neophodno je prisustvo ribozoma, aktivacija tRNK, pri čemu se vezuju odgovarajuće aminokiseline, prisustvo Mg2+ jona, kao i optimalni uslovi okoline (temperatura, pH, pritisak, itd.).
Za početak emitovanja ( iniciranje) mala ribosomalna podjedinica je vezana za molekul mRNA spreman za sintezu, a zatim se, prema principu komplementarnosti prvom kodonu (AUG), odabire tRNA koja nosi aminokiselinu metionin. Tek nakon toga se veže velika ribosomska podjedinica. Unutar sastavljenog ribozoma postoje dva kodona mRNA, od kojih je prvi već zauzet. Druga tRNA, koja također nosi aminokiselinu, dodaje se kodonu uz nju, nakon čega se uz pomoć enzima formira peptidna veza između aminokiselinskih ostataka. Ribosom pomiče jedan kodon mRNA; prva tRNA oslobođena aminokiseline vraća se u citoplazmu nakon sljedeće aminokiseline, a fragment budućeg polipeptidnog lanca visi, takoreći, na preostaloj tRNK. Sljedeća tRNA se veže za novi kodon koji se nađe unutar ribozoma, proces se ponavlja i korak po korak se polipeptidni lanac produžuje, tj. izduženje.
Kraj sinteze proteina ( prestanak) se javlja čim se u molekulu mRNA naiđe na specifičnu sekvencu nukleotida koja ne kodira aminokiselinu (stop kodon). Nakon toga se odvajaju ribosom, mRNA i polipeptidni lanac, a novosintetizovani protein dobija odgovarajuću strukturu i transportuje se u deo ćelije gde će obavljati svoje funkcije.
Translacija je energetski vrlo intenzivan proces, budući da se energija jedne molekule ATP-a troši na pričvršćivanje jedne aminokiseline na tRNA, a još nekoliko se koristi za pomicanje ribozoma duž mRNA molekula.
Da bi se ubrzala sinteza određenih proteinskih molekula, nekoliko ribozoma može se sukcesivno vezati za molekulu mRNA, koji formiraju jednu strukturu - polizom.
Ćelija je genetska jedinica živog bića. Kromosomi, njihova struktura (oblik i veličina) i funkcije. Broj hromozoma i konstantnost njihove vrste. Somatske i zametne ćelije. Životni ciklus ćelije: interfaza i mitoza. Mitoza je podjela somatskih ćelija. Mejoza. Faze mitoze i mejoze. Razvoj zametnih ćelija u biljkama i životinjama. Podjela ćelija je osnova za rast, razvoj i reprodukciju organizama. Uloga mejoze i mitoze
Ćelija je genetska jedinica živog bića.
Unatoč činjenici da su nukleinske kiseline nosilac genetske informacije, implementacija tih informacija je nemoguća izvan ćelije, što se lako dokazuje na primjeru virusa. Ovi organizmi, koji često sadrže samo DNK ili RNK, ne mogu se razmnožavati samostalno; da bi to učinili, moraju koristiti nasljedni aparat ćelije. Ne mogu čak ni prodrijeti u ćeliju bez pomoći same ćelije, osim upotrebom membranskih transportnih mehanizama ili zbog oštećenja ćelije. Većina virusa je nestabilna; umiru nakon samo nekoliko sati izlaganja na otvorenom. Shodno tome, ćelija je genetska jedinica živog bića, koja ima minimalan skup komponenti za očuvanje, promjenu i implementaciju nasljednih informacija, kao i njihovo prenošenje na potomke.
Većina genetskih informacija eukariotske ćelije nalazi se u jezgru. Posebnost njegove organizacije je da, za razliku od DNK prokariotske ćelije, molekuli DNK eukariota nisu zatvoreni i formiraju složene komplekse s proteinima - hromozomima.
Kromosomi, njihova struktura (oblik i veličina) i funkcije
hromozom(iz grčkog hrom- boja, boja i soma- tijelo) je struktura ćelijskog jezgra, koja sadrži gene i nosi određene nasljedne informacije o karakteristikama i svojstvima organizma.
Ponekad se kružni DNK molekuli prokariota nazivaju i hromozomi. Kromosomi su sposobni za samoumnožavanje, imaju strukturnu i funkcionalnu individualnost i zadržavaju je generacijama. Svaka ćelija nosi sve nasljedne informacije tijela, ali samo mali dio radi u njoj.
Osnova hromozoma je dvolančana DNK molekula prepuna proteina. Kod eukariota, histonski i nehistonski proteini stupaju u interakciju s DNK, dok kod prokariota histonski proteini odsutni.
Hromozomi se najbolje vide pod svjetlosnim mikroskopom tokom diobe ćelije, kada kao rezultat zbijanja poprimaju izgled štapićastih tijela razdvojenih primarnim suženjem - centromere — na ramenima. Može biti i na hromozomu sekundarna konstrikcija, koji u nekim slučajevima razdvaja tzv satelit. Zovu se krajevi hromozoma telomere. Telomeri sprečavaju da se krajevi hromozoma lepe zajedno i obezbeđuju njihovo pričvršćivanje za nuklearnu membranu u ćeliji koja se ne deli. Na početku diobe hromozomi se udvostručuju i sastoje se od dva ćerka hromozoma - hromatida, pričvršćen na centromeri.

Prema svom obliku, hromozomi se dijele na ravnokrake, nejednakokrake i štapićaste hromozome. Veličine hromozoma značajno variraju, ali prosječni hromozom ima dimenzije od 5 $×$ 1,4 mikrona.
U nekim slučajevima, hromozomi, kao rezultat brojnih duplikacija DNK, sadrže stotine i hiljade hromatida: takvi ogromni hromozomi se nazivaju polytene. Nalaze se u pljuvačnim žlijezdama larvi Drosophila, kao i u probavnim žlijezdama okruglih crva.
Broj hromozoma i konstantnost njihove vrste. Somatske i zametne ćelije
Prema ćelijskoj teoriji, ćelija je jedinica strukture, vitalne aktivnosti i razvoja organizma. Dakle, tako važne funkcije živih bića kao što su rast, reprodukcija i razvoj organizma su obezbeđene na ćelijskom nivou. Ćelije višećelijskih organizama mogu se podijeliti na somatske i reproduktivne ćelije.
Somatske ćelije- sve su to ćelije tijela nastale kao rezultat mitotičke diobe.
Proučavanje hromozoma omogućilo je da se utvrdi da se somatske ćelije tijela svake biološke vrste karakteriziraju konstantnim brojem kromosoma. Na primjer, osoba ih ima 46. Skup hromozoma somatskih ćelija se zove diploidni(2n), ili duplo.

Polne ćelije, ili gamete, su specijalizovane ćelije koje se koriste za seksualnu reprodukciju.
Gamete uvijek sadrže upola manje hromozoma od somatskih ćelija (kod ljudi - 23), pa se skup hromozoma zametnih ćelija naziva haploidni(n) ili pojedinac. Njegovo stvaranje povezano je s mejotskom diobom stanica.
Količina DNK u somatskim ćelijama označena je kao 2c, au polnim ćelijama - 1c. Genetska formula somatskih ćelija je zapisana kao 2n2c, a polnih ćelija - 1n1c.
U jezgrima nekih somatskih stanica broj kromosoma može se razlikovati od broja u somatskim stanicama. Ako je ta razlika veća od jednog, dva, tri, itd. haploidnih skupova, tada se takve ćelije nazivaju poliploid(tri-, tetra-, pentaploid, respektivno). U takvim ćelijama se metabolički procesi obično odvijaju vrlo intenzivno.
Broj hromozoma sam po sebi nije specifično obilježje vrste, budući da različiti organizmi mogu imati jednak broj hromozoma, ali srodni mogu imati različit broj. Na primjer, malarijski plazmodijum i konjski okrugli crv imaju po dva hromozoma, dok ljudi i čimpanze imaju 46, odnosno 48.
Ljudski hromozomi se dele u dve grupe: autozomi i polni hromozomi (heterohromozomi). Autosome u ljudskim somatskim ćelijama ima 22 para, isti su za muškarce i žene, i polni hromozomi samo jedan par, ali to je ono što određuje pol pojedinca. Postoje dvije vrste polnih hromozoma - X i Y. Ženske tjelesne ćelije nose dva X hromozoma, a muške - X i Y.
Kariotip- ovo je skup karakteristika hromozomskog seta organizma (broj kromosoma, njihov oblik i veličina).
Uslovni zapis kariotipa uključuje ukupan broj hromozoma, polnih hromozoma i moguća odstupanja u setu hromozoma. Na primjer, kariotip normalnog muškarca piše se kao 46, XY, a kariotip normalne žene je 46, XX.
Životni ciklus ćelije: interfaza i mitoza
Ćelije ne nastaju svaki put iznova, one nastaju samo kao rezultat diobe matičnih stanica. Nakon diobe ćelijama kćeri potrebno je neko vrijeme da formiraju organele i steknu odgovarajuću strukturu koja bi osigurala obavljanje određene funkcije. Ovaj vremenski period se zove sazrevanje.
Naziva se vremenski period od pojave ćelije kao rezultat diobe do njene diobe ili smrti životni ciklus ćelije.
U eukariotskim ćelijama životni ciklus je podijeljen u dvije glavne faze: interfazu i mitozu.

Interfaza- ovo je vremenski period u životnom ciklusu tokom kojeg se ćelija ne dijeli i normalno funkcionira. Interfaza je podijeljena na tri perioda: G 1 -, S- i G 2 -periodi.
G 1 -period(presintetički, postmitotski) je period rasta i razvoja ćelije tokom kojeg se odvija aktivna sinteza RNK, proteina i drugih supstanci neophodnih za potpuni život novonastale ćelije. Pred kraj ovog perioda, ćelija može početi da se priprema za dupliciranje svoje DNK.
IN S-period(sintetički) dolazi do samog procesa replikacije DNK. Jedini dio hromozoma koji ne podliježe replikaciji je centromera, pa se tako nastale DNK molekule ne divergiraju u potpunosti, već se drže zajedno u njemu, a na početku diobe hromozom ima X-oblik. Genetska formula ćelije nakon udvostručenja DNK je 2n4c. Takođe u S-periodu, centrioli ćelijskog centra su udvostručeni.
G 2 -period(postsintetski, premitotički) karakteriše intenzivna sinteza RNK, proteina i ATP-a neophodnih za proces deobe ćelije, kao i odvajanje centriola, mitohondrija i plastida. Do kraja interfaze, kromatin i nukleolus ostaju jasno razlučivi, integritet nuklearnog omotača nije narušen, a organele se ne mijenjaju.
Neke od tjelesnih stanica su u stanju da obavljaju svoje funkcije tokom cijelog života u tijelu (neuroni našeg mozga, mišićne ćelije srca), dok druge postoje kratko, nakon čega odumiru (ćelije crijevnog epitela, epidermalne ćelije koža). Shodno tome, tijelo mora stalno prolaziti kroz procese diobe stanica i formiranja novih koje bi zamijenile mrtve. Zovu se ćelije sposobne za dijeljenje stablo. U ljudskom tijelu nalaze se u crvenoj koštanoj srži, u dubokim slojevima epiderme kože i na drugim mjestima. Koristeći ove ćelije, možete uzgajati novi organ, postići podmlađivanje, a također i klonirati tijelo. Izgledi za korištenje matičnih stanica su apsolutno jasni, ali se još uvijek raspravlja o moralnim i etičkim aspektima ovog problema, jer se u većini slučajeva koriste embrionalne matične stanice dobivene iz ljudskih embriona ubijenih tijekom pobačaja.
Trajanje interfaze u biljnim i životinjskim ćelijama je u prosjeku 10-20 sati, dok mitoza traje oko 1-2 sata.
Tokom uzastopnih podjela u višećelijskim organizmima, ćelije kćeri postaju sve raznovrsnije jer čitaju informacije iz sve većeg broja gena.
Neke stanice se s vremenom prestaju dijeliti i umiru, što može biti zbog dovršetka određenih funkcija, kao u slučaju epidermalnih stanica kože i krvnih stanica, ili zbog oštećenja ovih stanica faktorima okoline, posebno patogenima. Genetski programirana ćelijska smrt se naziva apoptoza, dok nesretna smrt - nekroza.
Mitoza je podjela somatskih ćelija. Faze mitoze
Mitoza- metoda indirektne diobe somatskih stanica.
Tokom mitoze, ćelija prolazi kroz niz uzastopnih faza, usled čega svaka ćerka ćelija dobija isti set hromozoma kao i matična ćelija.
Mitoza je podijeljena u četiri glavne faze: profaza, metafaza, anafaza i telofaza. Profaza- najduža faza mitoze, tokom koje se hromatin kondenzuje, što rezultira hromozomima u obliku slova X koji se sastoje od dve hromatide (kromosoma kćeri) postaju vidljivi. U tom slučaju nukleolus nestaje, centriole se razilaze prema polovima stanice i počinje se formirati akromatinsko vreteno (diobno vreteno) iz mikrotubula. Na kraju profaze, nuklearna membrana se raspada u zasebne vezikule.

IN metafaza Kromosomi su poređani duž ekvatora ćelije sa svojim centromerama, za koje su pričvršćene mikrotubule potpuno formiranog vretena. U ovoj fazi podjele, kromosomi su najkompaktniji i imaju karakterističan oblik, što omogućava proučavanje kariotipa.
IN anafaza Brza replikacija DNK događa se na centromerama, zbog čega se hromozomi cijepaju i hromatide divergiraju do polova ćelije, rastegnute mikrotubulama. Raspodjela hromatida mora biti apsolutno jednaka, jer upravo ovaj proces osigurava održavanje konstantnog broja kromosoma u stanicama tijela.
Na sceni telofazećerki hromozomi se skupljaju na polovima, oko njih se iz vezikula formiraju despiralne, nuklearne membrane, a u novonastalim jezgrama pojavljuju se jezgre.
Nakon nuklearne diobe dolazi do diobe citoplazme - citokineza, tokom koje dolazi do manje ili više ujednačene raspodele svih organela matične ćelije.
Dakle, kao rezultat mitoze, iz jedne matične ćelije nastaju dvije kćerke ćelije, od kojih je svaka genetska kopija matične ćelije (2n2c).
U bolesnim, oštećenim, ostarjelim stanicama i specijaliziranim tkivima tijela može doći do nešto drugačijeg procesa diobe - amitoze. Amitoza naziva se direktna dioba eukariotskih stanica, u kojoj ne dolazi do formiranja genetski ekvivalentnih stanica, jer su ćelijske komponente neravnomjerno raspoređene. Nalazi se u biljkama u endospermu, a kod životinja - u jetri, hrskavici i rožnici oka.
Mejoza. Faze mejoze
Mejoza je metoda indirektne diobe primarnih zametnih stanica (2n2c), koja rezultira stvaranjem haploidnih stanica (1n1c), najčešće zametnih stanica.
Za razliku od mitoze, mejoza se sastoji od dvije uzastopne diobe stanica, od kojih svakoj prethodi interfaza. Prva podjela mejoze (mejoza I) se zove redukcionistički, budući da je u ovom slučaju broj hromozoma prepolovljen, a druga podjela (mejoza II) - equational, budući da je u njegovom procesu sačuvan broj hromozoma.

Interfaza I nastavlja kao interfaza mitoze. Mejoza I podijeljen je u četiri faze: profaza I, metafaza I, anafaza I i telofaza I. B profaza I Događaju se dva važna procesa: konjugacija i krosing. Konjugacija- Ovo je proces fuzije homolognih (uparenih) hromozoma cijelom dužinom. Parovi hromozoma nastali tokom konjugacije čuvaju se do kraja metafaze I.
Prelazak- međusobna izmjena homolognih regija homolognih hromozoma. Kao rezultat križanja, hromozomi koje tijelo primi od oba roditelja dobivaju nove kombinacije gena, što uzrokuje pojavu genetski raznolikog potomstva. Na kraju profaze I, kao iu profazi mitoze, jezgro nestaje, centriole se divergiraju prema polovima ćelije, a nuklearna membrana se raspada.

IN metafaza I parovi hromozoma su poredani duž ekvatora ćelije, a mikrotubule vretena su pričvršćene za njihove centromere.
IN anafaza I Cijeli homologni hromozomi, koji se sastoje od dvije hromatide, divergiraju do polova.
IN telofaza I Nuklearne membrane se formiraju oko klastera hromozoma na polovima ćelije i formiraju se jezgre.
Citokineza I osigurava odvajanje citoplazme ćelija kćeri.
Ćerke ćelije (1n2c) nastale kao rezultat mejoze I su genetski heterogene, jer njihovi hromozomi, nasumično raspoređeni na polove ćelije, sadrže različite gene.
Komparativne karakteristike mitoze i mejoze
| Potpiši | Mitoza | Mejoza | |
| Koje ćelije počinju da se dele? | somatski (2n) | Primarne zametne ćelije (2n) | |
| Broj divizija | 1 | 2 | |
| Koliko i kakvih ćelija se formira tokom deobe? | 2 somatske (2n) | 4 seksualne (n) | |
| Interfaza | Priprema ćelije za podelu, udvostručavanje DNK | Vrlo kratko, ne dolazi do udvostručavanja DNK | |
| Faze | Mejoza I | Mejoza II | |
| Profaza | Može doći do kondenzacije hromozoma, nestanka nukleola, dezintegracije nuklearne membrane, konjugacije i krosingovera | Kondenzacija hromozoma, nestanak nukleola, dezintegracija nuklearne membrane | |
| Metafaza | Parovi hromozoma nalaze se duž ekvatora, formira se vreteno | Hromozomi se nižu duž ekvatora, formira se vreteno | |
| Anafaza | Homologni hromozomi iz dve hromatide kreću se prema polovima | Kromatide se kreću prema polovima | |
| Telofaza | Kromosomi su despiralni, formiraju se nove nuklearne membrane i jezgre | Kromosomi su despiralni, formiraju se nove nuklearne membrane i jezgre | |
Interfaza II vrlo kratko, pošto u njemu ne dolazi do udvostručenja DNK, odnosno nema S-perioda.
Mejoza II također podijeljen u četiri faze: profaza II, metafaza II, anafaza II i telofaza II. IN profaza II isti procesi se dešavaju kao u profazi I, sa izuzetkom konjugacije i krosingovera.
IN metafaza II hromozomi se nalaze duž ekvatora ćelije.
IN anafaza II hromozomi su podijeljeni na centromerima i hromatide su rastegnute prema polovima.
IN telofaza II Nuklearne membrane i jezgre formiraju se oko klastera kćeri hromozoma.
Poslije citokineza II Genetska formula sve četiri ćelije kćeri je 1n1c, ali sve imaju drugačiji skup gena, što je rezultat ukrštanja i slučajne kombinacije hromozoma majčinih i očinskih organizama u ćelijama kćeri.
Razvoj zametnih ćelija u biljkama i životinjama
Gametogeneza(iz grčkog gameta- supruga, gamete- muž i geneza- nastanak, nastanak) je proces formiranja zrelih zametnih ćelija.
Budući da su za spolnu reprodukciju najčešće potrebne dvije individue - ženka i mužjak, koji proizvode različite spolne stanice - jajašca i spermu, onda se procesi formiranja ovih gameta moraju razlikovati.
Priroda procesa u značajnoj mjeri ovisi o tome da li se događa u biljnoj ili životinjskoj ćeliji, jer se kod biljaka javlja samo mitoza prilikom formiranja gameta, a kod životinja i mitoza i mejoza.
Razvoj zametnih ćelija u biljkama. Kod kritosjemenjača formiranje muških i ženskih reproduktivnih stanica događa se u različitim dijelovima cvijeta - prašnicima, odnosno tučkom.
Prije formiranja muških reproduktivnih stanica - mikrogametogeneza(iz grčkog mikros- mali) - dešava se mikrosporogeneza, odnosno stvaranje mikrospora u prašnicima prašnika. Ovaj proces je povezan s mejotičkom diobom matične stanice, što rezultira četiri haploidne mikrospore. Mikrogametogeneza je povezana s mitotskom diobom mikrospore, dajući muški gametofit iz dvije ćelije - velike vegetativno(sifonogena) i plitka generativno. Nakon diobe, muški gametofit postaje prekriven gustim membranama i formira polenovo zrno. U nekim slučajevima, čak i tokom procesa sazrevanja polena, a ponekad tek nakon prelaska na stigmu tučka, generativna ćelija se mitotički deli i formira dve nepokretne muške zametne ćelije - sperma. Nakon oprašivanja, iz vegetativne ćelije formira se polenova cijev kroz koju spermatozoidi prodiru u jajnik tučka radi oplodnje.

Razvoj ženskih zametnih ćelija u biljkama naziva se megagametogeneza(iz grčkog megas- veliki). Javlja se u jajniku tučka, kojem prethodi megasporogeneza, kao rezultat čega se mejotičkom diobom formiraju četiri megaspore iz matične stanice megaspore koja leži u nucelusu. Jedna od megaspora se mitotički dijeli tri puta, dajući ženskom gametofitu - embrionsku vrećicu sa osam jezgara. Uz naknadno odvajanje citoplazme ćelija kćeri, jedna od nastalih ćelija postaje jaje, na čijim stranama leže takozvani sinergidi, na suprotnom kraju embrionalne vrećice formiraju se tri antipoda, a u sredini , kao rezultat fuzije dva haploidna jezgra, nastaje diploidna centralna ćelija.
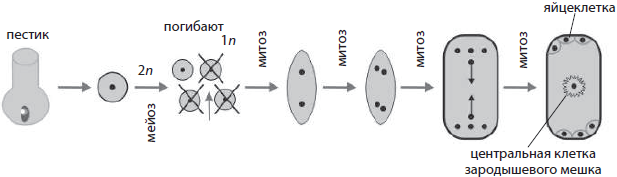
Razvoj zametnih ćelija kod životinja. Kod životinja postoje dva procesa formiranja zametnih ćelija - spermatogeneza i oogeneza.
Spermatogeneza(iz grčkog sperma, spermatozoidi- seme i geneza- porijeklo, pojava) je proces formiranja zrelih muških zametnih stanica - spermatozoida. Kod ljudi se javlja u testisima, odnosno testisima, i dijeli se na četiri perioda: razmnožavanje, rast, sazrijevanje i formiranje.

IN sezona razmnožavanja primordijalne zametne stanice dijele se mitotički, što rezultira stvaranjem diploida spermatogonije. IN period rasta spermatogonije akumuliraju hranjive tvari u citoplazmi, povećavaju se i pretvaraju u primarnih spermatocita, ili Spermatociti 1. reda. Tek nakon toga ulaze u mejozu ( period sazrevanja), kao rezultat čega se formiraju prva dva sekundarni spermatocit, ili Spermatocit 2. reda, a zatim - četiri haploidne ćelije sa još prilično velikom količinom citoplazme - spermatida. IN period formiranja gube gotovo svu svoju citoplazmu i formiraju flagellum, pretvarajući se u spermu.
Sperma, ili livelies, - vrlo male pokretne muške reproduktivne stanice s glavom, vratom i repom.

IN glava, pored jezgre, je akrozom- modificirani Golgi kompleks koji osigurava otapanje jajnih membrana tokom oplodnje. IN cerviksa su centrioli ćelijskog centra i baze konjski rep formiraju mikrotubule koje direktno podržavaju kretanje spermatozoida. Sadrži i mitohondrije, koje spermiju obezbjeđuju ATP energiju za kretanje.
Oogeneza(iz grčkog UN- jaje i geneza- porijeklo, pojava) je proces formiranja zrelih ženskih zametnih stanica - jaja. Kod ljudi se javlja u jajnicima i sastoji se od tri perioda: reprodukcije, rasta i sazrijevanja. Periodi razmnožavanja i rasta, slični onima u spermatogenezi, javljaju se tokom intrauterinog razvoja. U ovom slučaju, diploidne ćelije nastaju iz primarnih zametnih ćelija kao rezultat mitoze. oogonia, koji se zatim pretvaraju u diploidne primarne oociti, ili Oociti 1. reda. Mejoza i naknadna citokineza koja se javlja u period sazrevanja, karakteriše neravnomerna deoba citoplazme matične ćelije, tako da se kao rezultat toga dobija prvo sekundarne oocite, ili Oocita 2. reda, And prvo polarno tijelo, a zatim iz sekundarne oocite - jajeta, koje zadržava cjelokupnu zalihu hranjivih tvari, i drugo polarno tijelo, dok je prvo polarno tijelo podijeljeno na dva. Polarna tijela preuzimaju višak genetskog materijala.
Kod ljudi se jaja stvaraju u intervalu od 28-29 dana. Ciklus povezan sa sazrevanjem i oslobađanjem jajnih ćelija naziva se menstrualni.
Jaje- velika ženska reproduktivna stanica koja nosi ne samo haploidni skup hromozoma, već i značajnu zalihu hranjivih tvari za kasniji razvoj embrija.

Jaje kod sisara je prekriveno sa četiri membrane, koje smanjuju mogućnost oštećenja raznim faktorima. Promjer jajeta kod ljudi doseže 150-200 mikrona, dok u noja može biti nekoliko centimetara.
Podjela ćelija je osnova za rast, razvoj i reprodukciju organizama. Uloga mitoze i mejoze
Ako kod jednoćelijskih organizama dioba stanica dovodi do povećanja broja jedinki, odnosno reprodukcije, onda kod višećelijskih organizama ovaj proces može imati različita značenja. Dakle, podjela embrionalnih stanica, počevši od zigote, predstavlja biološka osnova međusobno povezanih procesa rasta i razvoja. Slične promjene se primjećuju kod ljudi tokom adolescencije, kada se broj ćelija ne samo povećava, već se javlja i kvalitativna promjena u tijelu. Razmnožavanje višećelijskih organizama zasniva se i na diobi ćelija, na primjer, kod aseksualne reprodukcije, zahvaljujući ovom procesu, obnavlja se cijeli dio organizma, a kod spolne reprodukcije u procesu gametogeneze nastaju polne ćelije koje nakon toga stvaraju novi organizam. Treba napomenuti da glavne metode diobe eukariotske ćelije - mitoza i mejoza - imaju različita značenja u životnim ciklusima organizama.
Kao rezultat mitoze, postoji ravnomjerna distribucija nasljednog materijala između stanica kćeri - točne kopije majke. Bez mitoze, postojanje i rast višećelijskih organizama koji se razvijaju iz jedne ćelije, zigote, bili bi nemogući, jer sve ćelije takvih organizama moraju sadržavati iste genetske informacije.
U procesu diobe ćelije kćeri postaju sve raznovrsnije po strukturi i funkcijama, što je povezano sa aktivacijom sve više novih grupa gena u njima zbog međustanične interakcije. Dakle, mitoza je neophodna za razvoj organizma.
Ova metoda diobe ćelija neophodna je za procese aseksualne reprodukcije i regeneracije (obnove) oštećenih tkiva, kao i organa.
Mejoza, zauzvrat, osigurava postojanost kariotipa tijekom seksualne reprodukcije, budući da prepolovi skup kromosoma prije seksualne reprodukcije, koji se zatim obnavlja kao rezultat oplodnje. Osim toga, mejoza dovodi do pojave novih kombinacija roditeljskih gena zbog križanja i nasumične kombinacije hromozoma u ćelijama kćerima. Zahvaljujući tome, potomci su genetski raznoliki, što daje materijal za prirodnu selekciju i predstavlja materijalnu osnovu za evoluciju. Promjena broja, oblika i veličine hromozoma, s jedne strane, može dovesti do pojave raznih odstupanja u razvoju organizma, pa čak i njegove smrti, as druge strane može dovesti do pojave jedinki. prilagođeniji okruženju.
Dakle, ćelija je jedinica rasta, razvoja i reprodukcije organizama.
§ 2. Glavne komponente eukariotske ćelije
Eukariotske ćelije (sl. 8 i 9) su organizovane mnogo složenije od prokariotskih. Vrlo su raznolike po veličini (od nekoliko mikrometara do nekoliko centimetara), po obliku i strukturnim karakteristikama (sl. 10).
Rice. 8. Struktura eukariotske ćelije. Generalizirana shema
Rice. 9. Struktura ćelije prema elektronskoj mikroskopiji
Rice. 10. Razne eukariotske ćelije: 1 – epitelne; 2 – krv (e – eritroitis, / – leukitis); 3 – hrskavica; 4 – kosti; 5 – glatki mišić; 6 – vezivno tkivo; 7 – nervne ćelije; 8 – prugasto mišićno vlakno
Međutim, opšta organizacija i prisustvo osnovnih komponenti su isti u svim eukariotskim ćelijama (slika 11).
Rice. 11. Eukariotska ćelija (dijagram)
Plazmalema (spoljna ćelijska membrana). Osnova plazma membrane, kao i drugih membrana u ćelijama (npr. mitohondrije, plastidi, itd.), je sloj lipida koji ima dva reda molekula (Sl. 12). Pošto su molekuli lipida polarni (jedan pol je hidrofilan, tj. privlači ga voda, a drugi je hidrofoban, tj. odbija se vodom), oni su raspoređeni u određenom redoslijedu. Hidrofilni krajevi molekula jednog sloja usmjereni su prema vodenoj sredini - u citoplazmu ćelije, a drugog sloja - prema van iz ćelije - prema međućelijskoj tvari (kod višećelijskih organizama) ili vodenoj sredini (kod jednoćelijskih organizama). ).
Rice. 12. Struktura ćelijske membrane prema modelu fluidnog mozaika. Proteini i glikoproteini su uronjeni u dvostruki sloj molekula lipida, sa svojim hidrofilnim krajevima (krugovima) okrenutim prema van, a hidrofobnim krajevima (valovitim linijama) okrenutim duboko u membranu.
Molekuli proteina su mozaično ugrađeni u bimolekularni sloj lipida. Na vanjskoj strani životinjske stanice, molekuli polisaharida su vezani za lipide i proteinske molekule plazmaleme, formirajući glikolipide i glikoproteine.
Ovaj agregat formira sloj glikokaliks. Povezan sa njim funkciju receptora plazma membrane (vidi dolje); takođe može akumulirati različite supstance koje koristi ćelija. Osim toga, glikokaliks poboljšava mehaničku stabilnost plazmaleme.
U stanicama biljaka i gljiva postoji i ćelijska stijenka koja ima potpornu i zaštitnu ulogu. U biljkama se sastoji od celuloze, a u gljivama od hitina.
Vanjska ćelijska membrana obavlja brojne funkcije, uključujući:
♦ mehanički(podrška, izgradnja forme);
♦ barijera-transport(selektivna propusnost za različite supstance: ulazak u ćeliju potrebnih supstanci i uklanjanje nepotrebnih i štetnih);
♦ receptor(određivanje različitih hemikalija koje se nalaze u neposrednoj blizini ćelije; percepcija signala u obliku hormona; prepoznavanje „stranog“ proteina od strane ćelija imunog sistema, itd.).
Razmjena tvari između ćelije i okoline odvija se na različite načine - pasivno i aktivno.
Molekule vode i raznih jona pasivno (zbog difuzije, osmoze), a da ćelija ne troši energiju, ulaze kroz posebne pore – to su pasivni transport. Makromolekule kao što su proteini, polisaharidi, čak i cijele ćelije, opskrbljuju se fagocitoza I pinocitoza sa potrošnjom energije – aktivni transport.
Fagocitozom se progutaju cijele stanice ili velike čestice (na primjer, razmislite o ishrani u amebama ili fagocitozi zaštitnih krvnih stanica bakterija). Tokom pinocitoze apsorbuju se male čestice ili kapljice tečne supstance. Zajedničko za oba procesa je da su apsorbirane tvari okružene invaginirajućom vanjskom membranom kako bi se formirala vakuola, koja se potom kreće duboko u citoplazmu ćelije.
Egzocitoza je proces (koji je takođe aktivan transport) u suprotnom smeru od fagocitoze i pinocitoze (slika 13). Uz njegovu pomoć mogu se ukloniti neprobavljeni ostaci hrane u protozoama ili biološki aktivne tvari nastale u sekretornoj stanici.
Citoplazma. Citoplazma je sadržaj ćelije, ograničen plazmalemom, sa izuzetkom jezgra. Sadrži osnovna tvar (hijaloplazma), organele I inkluzije.
Hyaloplasma- viskozna tečnost koja može biti u jednom i drugom stanju Zola(tečnost), ili gel(želeast).
Ako je potrebno, citoplazma je sposobna za reverzibilni prijelaz iz jednog stanja u drugo. Na primjer, tokom ameboidnog kretanja (zapamtite dio „Protozoe” iz kursa zoologije), tokom formiranja pseudopoda, dolazi do brzih prijelaza citoplazme iz gela u sol i obrnuto. To je zbog prisustva u citoplazmi velikog broja filamentoznih proteinskih molekula actina. Kada se međusobno povežu da formiraju trodimenzionalnu mrežu, citoplazma je u stanju gela, a kada se mreža raspadne, ona je u stanju sol.
Hijaloplazma sadrži različite tvari - enzime, proteine, ugljikohidrate, masti i druge, organske i mineralne. Ovdje se odvijaju različiti kemijski procesi – razgradnja tvari, njihova sinteza i modifikacije (promjene).
Organoidi. To su trajne komponente ćelije određene strukture i funkcije koje se nalaze u njenoj citoplazmi. U nastavku ćemo govoriti o organoidi opšte namene, svojstveno svim tipovima ćelija svih eukariota. Oni su povezani s osiguravanjem vitalnih funkcija potonjeg. Organoidi posebne namjene nalazi se samo u stanicama određenog (visoko specijaliziranog) tipa - na primjer, miofibrili u mišićnim stanicama.
Organele opšte namene imaju istu građu bez obzira na to kojim ćelijama i kojim organizmima pripadaju. Ali među njima postoje grupe sa membranom (endoplazmatski retikulum, Golgijev aparat, mitohondrije, plastidi, lizozomi, vakuole), kao i nemembranski ( ribosomi, ćelijski centar) struktura.
Endoplazmatski retikulum (ER). EPS se sastoji od membrana i predstavlja složen razgranati sistem tubula i cisterni koji prodire kroz celu citoplazmu ćelije (Sl. 14). Postoje dvije vrste EPS-a - grubo I glatko. Ribosomi su vezani za hrapave membrane (sa citoplazmatske strane), dok ih na glatkim membranama nema.
Rice. 14. Endoplazmatski retikulum
Endoplazmatski retikulum obavlja niz važnih funkcija u eukariotskoj ćeliji:
♦ razgraničenje(podjela unutrašnjeg volumena ćelije na različite reakcione prostore);
♦ učešće u sintezi organskih supstanci(ribozomi se nalaze na membranama grubog ER, a kompleksi enzima nalaze se na glatkim, osiguravajući sintezu lipida, ugljikohidrata itd.);
♦ učešće u formiranju elemenata Golgijevog aparata, lizosoma;
♦ transport supstanci.
Golgijev aparat. Golgijev aparat (AG) je sistem tenkovi(ravne vakuole) i mjehurići(vezikule), koje se nalaze u neposrednoj blizini jezgra ćelije, koje nastaju usled EPS-a kao rezultat odvajanja njegovih malih fragmenata (slika 15). Kada se ovi fragmenti spoje, nastaju nove cisterne Golgijevog aparata, dok se iz EPS-a transportuju različite supstance koje su uključene u sklapanje složenih organskih jedinjenja (proteini + ugljeni hidrati, proteini + lipidi itd.), koji se uklanjaju sa pomoć AG izvan ćelije. Ove biološki aktivne supstance se ili uklanjaju iz ćelije (koristeći sekretorne vakuole egzocitozom) ili su deo lizosoma (vidi dole) koje formira AG.
Rice. 15. Golgijev aparat:
Golgijev aparat obavlja sljedeće funkcije:
♦ sinteza biološki aktivne tvari koje proizvodi stanica;
♦ lučenje (uklanjanje iz ćelije) različitih supstanci(hormoni, enzimi, supstance od kojih je izgrađen ćelijski zid, itd.);
♦ učešće u formiranju lizosoma.
Mitohondrije. Svi tipovi eukariotskih ćelija imaju mitohondrije (slika 16). Izgledaju kao okrugla tijela ili šipke, rjeđe - niti. Njihove veličine se kreću od 1 do 7 mikrona. Broj mitohondrija u ćeliji kreće se od nekoliko stotina do desetina hiljada (kod velikih protozoa).
Rice. 16. Mitohondrije. Iznad - mitohondrije (a) u mokraćnim kanalima, vidljive u svjetlosnom mikroskopu. Ispod je trodimenzionalni model organizacije mitohondrija: 1 – kriste; 2 – vanjska membrana; 3 – unutrašnja membrana; 4 – matrica
Mitohondriju čine dvije membrane - vanjski I interni, između kojih se nalazi intermembranskog prostora. Unutrašnja membrana formira mnoge invaginacije - kriste, koje su ili ploče ili cijevi. Ova organizacija pruža ogromnu površinu unutrašnje membrane. Sadrži enzime koji osiguravaju pretvaranje energije sadržane u organskim tvarima (ugljikohidrati, lipidi) u ATP energiju, neophodnu za život stanice. Stoga je funkcija mitohondrija da učestvuje u energijećelijskih procesa. Zato je veliki broj mitohondrija svojstven, na primjer, mišićnim stanicama koje obavljaju mnogo posla.
Plastidi. U biljnim ćelijama nalaze se posebne organele - plastidi, koji često imaju vretenasti ili zaobljen oblik, ponekad složeniji. Postoje tri vrste plastida - hloroplasti (slika 17), hromoplasti i leukoplasti.
Hloroplasti razlikuju se po zelenoj boji, što je zbog pigmenta - hlorofil, osiguravanje procesa fotosinteza, odnosno sinteza organskih tvari iz vode (H 2 O) i ugljičnog dioksida (CO 2) korištenjem energije sunčeve svjetlosti. Kloroplasti se nalaze uglavnom u ćelijama lista (u višim biljkama). Sastoje se od dvije membrane koje se nalaze paralelno jedna s drugom i okružuju sadržaj hloroplasta - stroma. Unutrašnja membrana formira brojne spljoštene vrećice - tilakoidi, koji su naslagani (kao hrpa novčića) – žitarice - i leže u stromi. To su tilakoidi koji sadrže hlorofil.
Hromoplasti određuju žutu, narandžastu i crvenu boju mnogih cvjetova i plodova, u čijim ćelijama su prisutni u velikim količinama. Glavni pigmenti u njihovom sastavu su karoteni. Funkcionalna svrha kromoplasta je da privuče životinje bojom, osiguravajući oprašivanje cvijeća i raspršivanje sjemena.
Rice. 17. Plastidi: a – hloroplasti u ćelijama lista Elodea, vidljivi u svetlosnom mikroskopu; b – dijagram unutrašnje strukture hloroplasta sa granama, koje su hrpe ravnih vrećica koje se nalaze okomito na površinu hloroplasta; c – detaljniji dijagram koji prikazuje anastomozne cijevi koje povezuju pojedinačne komore ventilatora
Leukoplasti su bezbojni plastidi sadržani u ćelijama podzemnih dijelova biljaka (na primjer, u gomoljima krumpira), sjemenkama i jezgri stabljike. U leukoplastima škrob se uglavnom formira iz glukoze i akumulira u organima za skladištenje biljaka.
Plastidi jedne vrste mogu se transformirati u drugu. Na primjer, kada lišće promijeni boju u jesen, hloroplasti se pretvaraju u hromoplaste.
Lizozomi. Ove organele izgledaju kao vezikule okružene membranom promjera do 2 mikrona. Sadrže nekoliko desetina enzima koji razgrađuju proteine, nukleinske kiseline, polisaharide i lipide. Funkcija lizosoma je da učestvuju u procesima unutarstanične razgradnje složenih organskih jedinjenja (na primer, hranljivih materija ili supstanci „potrošenih“ ćelijskih komponenti). Lizosomi se spajaju s fagocitnim (ili pinocitnim) vakuolama i formiraju digestivnu vakuolu.
Do stvaranja lizosoma dolazi zbog pupanja iz cisterni Golgijevog aparata.
Ribosomi. Ribosomi (slika 18) su prisutni u ćelijama i eukariota i prokariota, budući da obavljaju važnu funkciju u biosinteza proteina(vidi poglavlje 5). Svaka ćelija sadrži desetine, stotine hiljada (do nekoliko miliona) ovih malih okruglih organela.
Rice. 18. Šema strukture ribozoma koji se nalazi na membrani endoplazmatskog retikuluma: 1 – mala podjedinica; 2 – tRNA; 3 – aminoacil-tRNA; 4 – aminokiselina; 5 – velika podjedinica; 6 – membrana endoplazmatskog retikuluma; 7 – sintetizovani polipeptidni lanac
Ribosom se sastoji od dvije nejednake podjedinice (dijela). Oni se formiraju odvojeno i kombinuju, "obuhvatajući" glasničku RNK, tokom sinteze proteinskog molekula. Ribosomi sadrže različite proteine i ribosomske RNK.
Ćelijske inkluzije. Ovo je naziv za nestalne komponente u ćeliji, prisutne u glavnoj supstanci citoplazme u obliku zrnaca, granula ili kapljica. Inkluzije mogu ili ne moraju biti okružene membranom.
Funkcionalno, postoje tri vrste inkluzija: rezerve hranljivih materija(skrob, glikogen, masti, proteini), sekretorne inkluzije(supstance karakteristične za ćelije žlezda, koje one proizvode - hormoni endokrinih žlezda itd.) i uključivanje posebne namjene(u visokospecijaliziranim stanicama, na primjer hemoglobin u crvenim krvnim zrncima).
§ 3. Organizacija ćelijskog jezgra. hromozomi
Ćelijsko jezgro (vidi slike 8 i 9) je od najveće važnosti u životu ćelije, jer služi kao skladište naslednih informacija sadržanih u hromozomima (vidi dole).
Jezgro je ograničeno nuklearnim omotačem koji odvaja njegov sadržaj (karioplazma) iz citoplazme. Školjka se sastoji od dvije membrane koje su međusobno razdvojene. Oba su prožeta brojnim porama, zahvaljujući kojima je moguća izmjena tvari između jezgre i citoplazme. U ćelijskom jezgru većine eukariota ima od 1 do 7 nucleoli. S njima su povezani procesi sinteze RNK i tRNA.
Glavne komponente kernela – hromozomi, nastala od molekula DNK i raznih proteina. U svjetlosnom mikroskopu jasno su vidljivi samo u periodu diobe ćelije (mitoza, mejoza). U ćeliji koja se ne dijeli, hromozomi izgledaju kao dugačke tanke niti raspoređene po cijelom volumenu jezgra.
Tokom diobe ćelije, hromozomske niti formiraju guste spirale, zbog čega postaju vidljive (koristeći konvencionalni mikroskop) u obliku štapića, "ukosnica". Cjelokupna količina genetskih informacija je raspoređena između hromozoma jezgra. U procesu njihovog proučavanja identifikovani su sljedeći obrasci:
♦ u jezgrima somatskih ćelija (tj. tjelesnih ćelija, neseksualnih) sve jedinke iste vrste sadrže isti broj hromozoma koji čine skup hromozoma(Sl. 19);
Rice. 19. Hromozomi različitih vrsta biljaka i životinja, prikazani u istoj mjeri: 1,2 – ameba; 3,4 – dijatomeje; 5–8, 18,19 – zelene alge; 9 – muharica; 10 – lipa; 11–12 – Drosophila; 13 – losos; 14 – skerda (familija Asteraceae); 15 – biljka iz porodice aroidnih; 16 – leptir corydalis; 17 – insekt iz porodice skakavaca; 20 – buba vodoskoka; 21 – cvjetna buba; 22 – amfibijski ambistoma; 23 – aloja (porodica ljiljana)
♦ svaka vrsta se odlikuje vlastitim hromozomskim skupom prema broju (na primjer, osoba ima 46 hromozoma, muva Drosophila ima 8, okrugli crv ima 4, rak ima 196, konj ima 66, a kukuruz ima 104);
♦ hromozomi u jezgrama somatskih ćelija mogu se grupirati u parove tzv homolognih hromozoma na osnovu njihove sličnosti (u strukturi i funkciji);
♦ u jezgrima zametnih ćelija (gamete) svaki par homolognih hromozoma sadrži samo jedan, tj. ukupan skup hromozoma je upola manji od somatskih ćelija;
♦ jedan skup hromozoma u zametnim ćelijama naziva se haploidni i označava se slovom n, a u somatskim - diploidni(2n).
Iz navedenog je jasno da svaki par homolognih hromozoma nastaje spajanjem očevih i majčinih hromozoma tokom oplodnje, odnosno fuzijom polnih ćelija (gameta). Nasuprot tome, tokom formiranja zametnih ćelija, iz svakog para homolognih hromozoma samo jedan ulazi u gametu.
hromozomi drugačije homologni parovi se razlikuju po veličini i obliku (sl. 20 i 21).
Rice. 20. Struktura i tipovi hromozoma: a – izgled 1 – centromera; 2 – kratko rame; 3 – dugo rame); unutrašnja struktura istog hromozoma (1 – centromera; – DNK molekuli); c – tipovi hromozoma (1 – jednokraki; različitokraki; 3 – jednakokraki: X – krak, V – centromera)
Rice. 21. Hromozom se sastoji od DNK i proteina. Molekul DNK se replicira. Dvije identične dvostruke spirale DNK ostaju povezane u području centromera. Ove kopije postaju odvojeni hromozomi kasnije tokom ćelijske deobe
U tijelu postoje hromozomi primarna konstrikcija (nazvana centromera), na koji su konci pričvršćeni fisiona vretena. Ona dijeli hromozom na dva ramena Hromozomi mogu biti jednakoruki, nejednakoruki ili jednoruki.
Poglavlje 5. Metabolizam
§ 1. Metabolizam kao jedinstvo asimilacije i disimilacije
Sve ćelije i živi organizmi su otvoreni sistemi, odnosno u stanju su stalne razmene energija i supstanci sa okolinom. U neživoj prirodi postoje otvoreni sistemi, ali njihovo postojanje se kvalitativno razlikuje od živih organizama. Razmotrimo ovaj primjer: zapaljeni komad prirodnog sumpora je u stanju razmjene sa okolinom. Kada sagorijeva, O 2 se apsorbira, a SO 2 i energija (u obliku topline) se oslobađaju. Međutim, u ovom slučaju, komad sumpora kao fizičkog tijela se uništava i gubi svoju primarnu strukturu.
Za žive organizme, razmena sa okolinom se pokazuje kao uslov za očuvanje i održavanje njihove strukturne organizacije kroz samoobnavljanje svih supstanci i komponenti od kojih se sastoje.
Metabolizam (metabolizam) je skup procesa koji se odvijaju u živim organizmima (potrošnja, transformacija, akumulacija i oslobađanje tvari i energije) koji osiguravaju njihovu vitalnu aktivnost, razvoj, rast, reprodukciju. Tokom metaboličkog procesa, molekuli koji čine ćelije se razgrađuju i sintetišu; obnavljanje ćelijskih struktura i međućelijske supstance.
Metabolizam se zasniva na međusobno povezanim procesima asimilacija(anabolizam) i disimilacija(katabolizam). Tokom asimilacije (plastične razmjene), složene tvari se sintetiziraju iz jednostavnih. Zahvaljujući tome nastaju sve organske supstance u ćeliji neophodne za izgradnju njenih strukturnih komponenti, enzimskih sistema itd. Asimilacija se uvijek odvija uz trošenje energije.
Tijekom disimilacije (energetski metabolizam) složene organske tvari se razlažu na jednostavnije ili neorganske. U tom slučaju se oslobađa energija koju ćelija troši za obavljanje različitih procesa koji osiguravaju njenu vitalnu aktivnost (sinteza i transport tvari, mehanički rad, itd.).
Svi živi organizmi mogu se podijeliti u dvije grupe: autotrofi I heterotrofi, koje se razlikuju po izvorima energije i potrebnim supstancama za osiguranje svojih vitalnih funkcija.
Autotrofi– organizmi koji sintetiziraju organska jedinjenja iz neorganskih supstanci koristeći energiju sunčeve svjetlosti (npr fototrofi– biljke, cijanobakterije) ili energija dobivena oksidacijom mineralnih (anorganskih) tvari (npr. hemotrofi– bakterije sumpora, bakterije gvožđa itd.). Shodno tome, oni su u stanju samostalno stvarati tvari potrebne za njihovu životnu aktivnost.
§ 2. Disimilacija u anaerobnim i aerobnim organizmima
Organizmi se mogu podijeliti u dvije grupe i prema prirodi disimilacije - aerobes I anaerobi. Aerobes (od grč. demon– vazduh) zahtevaju slobodan kiseonik za život. Kod anaerobnih (grč. ac– negativna čestica) nije potrebno. U njima se fermentacijom vrši disimilacija - enzimska razgradnja organske tvari bez kisika uz stvaranje jednostavnijih organskih tvari i oslobađanje energije. Na primjer:
♦ fermentacija mliječne kiseline:
C 6 H 12 O 6 + 2H 3 PO 4 + 2ADP → 2PH + 2ATP + 2H 2 O;
♦ alkoholna fermentacija:
C 6 H 12 O 6 + 2PH + 2ADP → 2C 2 H 5 OH + 2CO 2 + 2ATP + 2H 2 O.
Tvari koje nastaju tokom fermentacije su organske i stoga još uvijek sadrže mnogo energije.
Rice. 22. Odnosi između asimilacije i disimilacije kod autotrofnih i heterotrofnih organizama
Kod aerobnih organizama tokom procesa disanja u mitohondrijima dolazi do potpunog razlaganja organskih materija (koristeći O 2) do energetski siromašnih krajnjih proizvoda CO 2 i H 2 O i oslobađa se znatno veća količina energije:
C 6 H 12 0 6 (glukoza) + 0 2 > 6C0 2 + 6H 2 0 + energija (zbog koje se sintetiše 38 ATP molekula).
Razmotrimo u obliku generaliziranih shema metabolizam autotrofnih i heterotrofnih aerobnih organizama (slika 22).
Asimilacija. Njegovi najvažniji procesi su fotosinteza I biosinteza proteina.
Razmjena supstanci koje ulaze u ćeliju ili se iz nje oslobađaju van, kao i razmjena različitih signala sa mikro- i makrookruženjem, odvija se kroz vanjsku membranu ćelije. Kao što je poznato, ćelijska membrana je lipidni dvosloj u koji su ugrađeni različiti proteinski molekuli koji djeluju kao specijalizirani receptori, jonski kanali, uređaji koji aktivno transportuju ili uklanjaju različite kemikalije, međućelijski kontakti itd. U zdravim eukariotskim ćelijama fosfolipidi su raspoređeni u membrana asimetrično: vanjska površina se sastoji od sfingomijelina i fosfatidilholina, unutrašnja površina - od fosfatidilserina i fosfatidiletanolamina. Održavanje takve asimetrije zahtijeva utrošak energije. Stoga se u slučaju oštećenja ćelije, infekcije ili energetskog gladovanja vanjska površina membrane obogaćuje za nju neuobičajenim fosfolipidima, što postaje signal drugim stanicama i enzimima o oštećenju stanice uz odgovarajuću reakciju na to. Najvažniju ulogu ima rastvorljivi oblik fosfolipaze A2, koji razgrađuje arahidonsku kiselinu i stvara lizoforme od gore navedenih fosfolipida. Arahidonska kiselina je ograničavajuća karika za stvaranje inflamatornih medijatora kao što su eikozanoidi, a zaštitni molekuli - pentraksini (C-reaktivni protein (CRP), prekursori amiloidnih proteina) - su vezani za lizoforme u membrani, nakon čega slijedi aktivacija komplementa. sistema duž klasičnog puta i destrukcije ćelija.
Struktura membrane pomaže u očuvanju karakteristika unutrašnjeg okruženja ćelije, njenih razlika od spoljašnjeg okruženja. To je osigurano selektivnom propusnošću ćelijske membrane i postojanjem aktivnih transportnih mehanizama u njoj. Njihov poremećaj kao rezultat direktnog oštećenja, na primjer, tetrodotoksinom, ouabainom, tetraetilamonijem, ili u slučaju nedovoljne opskrbe energijom odgovarajućih „pumpi“ dovodi do narušavanja sastava elektrolita ćelije, promjena u njenom metabolizmu, poremećaja. specifičnih funkcija – kontrakcija, provođenje pobudnih impulsa itd. Poremećaj ćelijskih jonskih kanala (kalcijum, natrijum, kalijum i hlorid) kod ljudi takođe može biti genetski uslovljen mutacijom gena odgovornih za strukturu ovih kanala. Takozvane kanalopatije uzrokuju nasljedne bolesti nervnog, mišićnog i probavnog sistema. Prekomjeran ulazak vode u ćeliju može dovesti do njenog pucanja – citolize – zbog perforacije membrane pri aktiviranju komplementa ili napada citotoksičnih limfocita i prirodnih stanica ubojica.
Stanična membrana ima mnogo receptora ugrađenih u nju - strukture koje, u kombinaciji s odgovarajućim specifičnim signalnim molekulima (ligandima), prenose signal unutar ćelije. To se događa kroz različite regulatorne kaskade koje se sastoje od enzimatski aktivnih molekula koji se sekvencijalno aktiviraju i na kraju doprinose implementaciji različitih ćelijskih programa, kao što su rast i proliferacija, diferencijacija, pokretljivost, starenje i ćelijska smrt. Regulatorne kaskade su prilično brojne, ali njihov broj još nije u potpunosti utvrđen. Sistem receptora i regulatornih kaskada povezanih sa njima takođe postoji unutar ćelije; stvaraju specifičnu regulatornu mrežu sa tačkama koncentracije, distribucije i odabira daljeg signalnog puta u zavisnosti od funkcionalnog stanja ćelije, faze njenog razvoja i istovremenog delovanja signala sa drugih receptora. Rezultat toga može biti inhibicija ili jačanje signala, usmjeravajući ga duž drugačijeg regulatornog puta. I receptorski aparat i putevi transdukcije signala kroz regulatorne kaskade, na primjer do jedra, mogu biti poremećeni kao rezultat genetskog defekta koji se javlja kao urođeni defekt na nivou organizma ili zbog somatske mutacije u specifičnom tipu ćelije. Ovi mehanizmi mogu biti oštećeni infektivnim agensima, toksinima, a mogu se promijeniti i tokom procesa starenja. Posljednja faza ovoga može biti poremećaj funkcija ćelije, procesa njezine proliferacije i diferencijacije.
Na površini ćelija nalaze se i molekuli koji igraju važnu ulogu u procesima međustanične interakcije. To mogu uključivati proteine stanične adhezije, antigene histokompatibilnosti, tkivno-specifične, diferencirajuće antigene itd. Promjene u sastavu ovih molekula uzrokuju poremećaj međućelijskih interakcija i mogu uzrokovati aktivaciju odgovarajućih mehanizama za eliminaciju takvih stanica, jer predstavljaju određene opasnosti po integritet organizma kao rezervoara infekcije, posebno virusne, ili kao potencijalnih pokretača rasta tumora.
Kršenje opskrbe energijom ćelije
Izvor energije u ćeliji je hrana, nakon čijeg se razgradnje energija oslobađa u konačne supstance. Glavno mjesto proizvodnje energije su mitohondrije, u kojima se tvari oksidiraju uz pomoć enzima respiratornog lanca. Oksidacija je glavni dobavljač energije, jer se kao rezultat glikolize ne oslobađa više od 5% energije iz iste količine oksidacijskih supstrata (glukoze), u odnosu na oksidaciju. Oko 60% energije koja se oslobađa tokom oksidacije akumulira se oksidativnom fosforilacijom u visokoenergetskim fosfatima (ATP, kreatin fosfat), a ostatak se raspršuje kao toplota. Nakon toga, visokoenergetske fosfate koristi ćelija za procese kao što su rad pumpe, sinteza, dioba, kretanje, izlučivanje itd. Postoje tri mehanizma čija oštećenja mogu uzrokovati poremećaj u opskrbi ćelijske energije: prvi je mehanizam sinteze enzima energetskog metabolizma, drugi je mehanizam oksidativne fosforilacije, treći je mehanizam korištenja energije.
Poremećaj transporta elektrona u mitohondrijskom respiratornom lancu ili razdvajanje ADP oksidacije i fosforilacije sa gubitkom protonskog potencijala, pokretačke sile za stvaranje ATP-a, dovodi do slabljenja oksidativne fosforilacije na način da se većina energije raspršuje kao toplota i smanjuje se broj visokoenergetskih spojeva. Razdvajanje oksidacije i fosforilacije pod uticajem adrenalina koriste ćelije homeotermnih organizama za povećanje proizvodnje toplote uz održavanje konstantne telesne temperature tokom hlađenja ili povećanje tokom groznice. Kod tireotoksikoze se uočavaju značajne promjene u strukturi mitohondrija i energetskom metabolizmu. Ove promjene su u početku reverzibilne, ali nakon određenog trenutka postaju ireverzibilne: mitohondrije se fragmentiraju, raspadaju ili nabubre, gube kriste, pretvaraju se u vakuole i na kraju akumuliraju tvari kao što su hijalin, feritin, kalcij, lipofuscin. Kod pacijenata sa skorbutom, mitohondrije se spajaju i formiraju hondriosfere, vjerovatno zbog oštećenja membrane peroksidnim spojevima. Značajna oštećenja mitohondrija nastaju pod uticajem jonizujućeg zračenja prilikom transformacije normalne ćelije u malignu.
Mitohondrije su moćan depo jona kalcijuma, gde je njegova koncentracija nekoliko redova veličine veća od one u citoplazmi. Kada su mitohondriji oštećeni, kalcij ulazi u citoplazmu, uzrokujući aktivaciju proteinaza s oštećenjem intracelularnih struktura i poremećajem funkcija odgovarajuće stanice, na primjer, kalcijeve kontrakture ili čak "kalcijevu smrt" u neuronima. Kao rezultat narušavanja funkcionalnog kapaciteta mitohondrija, naglo se povećava stvaranje slobodnih radikala peroksidnih spojeva, koji imaju vrlo visoku reaktivnost i stoga oštećuju važne komponente stanice - nukleinske kiseline, proteine i lipide. Ova pojava se uočava tokom takozvanog oksidativnog stresa i može imati negativne posledice na postojanje ćelije. Dakle, oštećenje vanjske membrane mitohondrija je praćeno oslobađanjem u citoplazmu tvari sadržanih u međumembranskom prostoru, prvenstveno citokroma C i nekih drugih biološki aktivnih tvari, koje pokreću lančane reakcije koje uzrokuju programiranu smrt stanice – apoptozu. Oštećujući mitohondrijsku DNK, reakcije slobodnih radikala iskrivljuju genetske informacije neophodne za formiranje određenih enzima respiratornog lanca, koji se proizvode posebno u mitohondrijima. To dovodi do još većeg poremećaja oksidativnih procesa. Generalno, sopstveni genetski aparat mitohondrija, u poređenju sa genetskim aparatom jezgra, manje je zaštićen od štetnih uticaja koji mogu promeniti genetske informacije koje su u njemu kodirane. Kao rezultat toga, disfunkcija mitohondrija se javlja tijekom cijelog života, na primjer, tokom procesa starenja, tokom maligne transformacije ćelije, kao iu pozadini nasljednih mitohondrijalnih bolesti povezanih s mutacijom mitohondrijske DNK u jajetu. Trenutno je opisano preko 50 mitohondrijalnih mutacija koje uzrokuju nasljedne degenerativne bolesti nervnog i mišićnog sistema. Prenose se na dijete isključivo od majke, jer mitohondrije sperme nisu dio zigota i, shodno tome, novog organizma.
Kršenje očuvanja i prijenosa genetskih informacija
Ćelijsko jezgro sadrži većinu genetskih informacija i na taj način osigurava njegovo normalno funkcioniranje. Kroz selektivnu ekspresiju gena, koordinira ćelijsku aktivnost tokom interfaze, pohranjuje genetske informacije i rekreira i prenosi genetski materijal tokom ćelijske diobe. Replikacija DNK i transkripcija RNK se dešavaju u jezgru. Razni patogeni faktori, kao što su ultraljubičasto i jonizujuće zračenje, oksidacija slobodnih radikala, hemikalije, virusi, mogu oštetiti DNK. Procjenjuje se da svakoj ćeliji toplokrvne životinje treba 1 dan. gubi više od 10.000 baza. Ovdje treba dodati prekršaje prilikom kopiranja tokom podjele. Ako se ova oštećenja nastave, ćelija ne bi mogla preživjeti. Zaštita leži u postojanju moćnih sistema popravke, kao što je ultraljubičasta endonukleaza, replikacija replikacije i rekombinacioni sistemi popravke, koji zamenjuju oštećenje DNK. Genetski defekti sistema popravke uzrokuju razvoj bolesti uzrokovanih povećanom osjetljivošću na faktore koji oštećuju DNK. To je pigmentna kseroderma, kao i neki sindromi ubrzanog starenja, praćeni povećanom tendencijom razvoja malignih tumora.
Sistem za regulisanje procesa replikacije DNK, transkripcije glasničke RNK (mRNA) i prevođenja genetskih informacija iz nukleinskih kiselina u strukturu proteina je prilično složen i višestepen. Pored regulatornih kaskada koje pokreću djelovanje transkripcionih faktora sa ukupnim brojem od preko 3000, koji aktiviraju određene gene, postoji i višeslojni regulatorni sistem posredovan malim RNK molekulima (interferirajuća RNA; RNAi). Ljudski genom, koji se sastoji od približno 3 milijarde purinskih i pirimidinskih baza, sadrži samo 2% strukturnih gena odgovornih za sintezu proteina. Ostatak obezbeđuje sintezu regulatornih RNK, koje istovremeno sa transkripcionim faktorima aktiviraju ili blokiraju rad strukturnih gena na nivou DNK u hromozomima ili utiču na procese translacije glasničke RNK (mRNA) tokom formiranja polipeptidne molekule u hromozomima. citoplazma. Povreda genetskih informacija može se javiti kako na nivou strukturnih gena, tako i na regulatornom dijelu DNK sa odgovarajućim manifestacijama u obliku različitih nasljednih bolesti.
U posljednje vrijeme veliku pažnju privlače promjene u genetskom materijalu koje nastaju tijekom individualnog razvoja organizma, a povezane su sa inhibicijom ili aktivacijom određenih dijelova DNK i hromozoma zbog njihove metilacije, acetilacije i fosforilacije. Ove promjene traju dugo, ponekad i tokom cijelog života organizma od embriogeneze do starosti, a nazivaju se epigenomskim naslijeđem.
Proliferaciju ćelija sa izmenjenim genetskim informacijama takođe sprečavaju sistemi (faktori) koji kontrolišu mitotički ciklus. Oni stupaju u interakciju s protein kinazama zavisnim od ciklina i njihovim katalitičkim podjedinicama - ciklinima - i blokiraju ćeliju da prođe kroz puni mitotički ciklus, zaustavljajući podjelu na granici između presintetičke i sintetičke faze (G1/S blok) dok se popravak DNK ne završi, a ako je to nemoguće, oni pokreću programirane ćelije smrti. Ovi faktori uključuju p53 gen, čija mutacija uzrokuje gubitak kontrole nad proliferacijom transformiranih ćelija; primećuje se u skoro 50% slučajeva raka kod ljudi. Druga kontrolna tačka mitotičkog ciklusa je na granici G2/M. Ovdje se ispravna distribucija hromozomskog materijala između ćelija kćeri u mitozi ili mejozi kontrolira korištenjem skupa mehanizama koji kontroliraju ćelijsko vreteno, centar i centromere (kinetohore). Neefikasnost ovih mehanizama dovodi do narušavanja distribucije hromozoma ili njihovih delova, što se manifestuje odsustvom bilo kog hromozoma u jednoj od ćelija kćeri (aneuploidija), prisustvom dodatnog hromozoma (poliploidija), odvajanjem hromozoma. dio hromozoma (delecija) i njegov prijenos na drugi hromozom (translokacija). Ovakvi procesi se vrlo često uočavaju prilikom proliferacije maligno degenerisanih i transformisanih ćelija. Ako se to dogodi tokom mejoze sa zametnim stanicama, to dovodi ili do smrti fetusa u ranoj fazi embrionalnog razvoja, ili do rođenja organizma s hromozomskom bolešću.
Nekontrolisana proliferacija ćelija tokom rasta tumora nastaje kao rezultat mutacija u genima koji kontrolišu proliferaciju ćelija i nazivaju se onkogeni. Među više od 70 trenutno poznatih onkogena, većina pripada komponentama regulacije ćelijskog rasta, neki su predstavljeni faktorima transkripcije koji regulišu aktivnost gena, kao i faktorima koji inhibiraju diobu i rast ćelija. Drugi faktor koji ograničava prekomjerno širenje (širenje) proliferirajućih stanica je skraćivanje krajeva hromozoma – telomera, koji se ne mogu u potpunosti replicirati kao rezultat čisto sterične interakcije, pa se nakon svake diobe ćelije telomeri skraćuju za određeni dio baza. Tako proliferirajuće ćelije odraslog organizma nakon određenog broja dioba (obično od 20 do 100 ovisno o vrsti organizma i njegovoj starosti) iscrpljuju dužinu telomera i zaustavlja se daljnja replikacija hromozoma. Ova pojava se ne javlja u epitelu spermatozoida, enterocitima i embrionalnim ćelijama zbog prisustva enzima telomeraze, koji vraća dužinu telomera nakon svake diobe. U većini ćelija odraslih organizama telomeraza je blokirana, ali se, nažalost, aktivira u tumorskim ćelijama.
Veza između jezgra i citoplazme i transport tvari u oba smjera odvija se kroz pore u nuklearnoj membrani uz sudjelovanje posebnih transportnih sistema koji troše energiju. Na taj način se energija i plastične tvari, signalni molekuli (transkripcijski faktori) transportuju do jezgra. Obrnuti tok nosi u citoplazmu molekule mRNA i transfer RNK (tRNA), ribozome neophodne za sintezu proteina u ćeliji. Isti put transporta supstanci je svojstven i virusima, posebno kao što je HIV. Oni prenose svoj genetski materijal u jezgro ćelije domaćina sa njegovom daljnjom inkorporacijom u genom domaćina i prijenosom novonastale virusne RNK u citoplazmu radi dalje sinteze proteina novih virusnih čestica.
Kršenje procesa sinteze
Procesi sinteze proteina odvijaju se u cisternama endoplazmatskog retikuluma, usko povezanim s porama u nuklearnoj membrani, kroz koje ribosomi, tRNA i mRNA ulaze u endoplazmatski retikulum. Ovdje se vrši sinteza polipeptidnih lanaca, koji naknadno dobijaju svoj konačni oblik u agranularnom endoplazmatskom retikulumu i lamelarnom kompleksu (Golgijev kompleks), gdje se podvrgavaju posttranslacijskoj modifikaciji i spajaju s molekulama ugljikohidrata i lipida. Novoformirani proteinski molekuli ne ostaju na mjestu sinteze, već kroz složen regulirani proces tzv. proteinkineza, aktivno se prenose u onaj izolirani dio ćelije gdje će obavljati svoju predviđenu funkciju. U ovom slučaju, vrlo važan korak je strukturiranje prenesene molekule u odgovarajuću prostornu konfiguraciju koja može obavljati svoju inherentnu funkciju. Ovo strukturiranje se dešava uz pomoć posebnih enzima ili na matrici specijalizovanih proteinskih molekula - šaperona, koji pomažu da molekul proteina, novoformiran ili promenjen usled spoljašnjeg uticaja, dobije ispravnu trodimenzionalnu strukturu. U slučaju štetnog djelovanja na ćeliju, kada postoji mogućnost narušavanja strukture proteinskih molekula (na primjer, povećanjem tjelesne temperature, infektivnim procesom, intoksikacijom), koncentracija pratioca u ćeliji se povećava. oštro. Stoga se takvi molekuli još nazivaju proteini stresa, ili proteini toplotnog šoka. Kršenje strukture proteinske molekule dovodi do stvaranja hemijski inertnih konglomerata, koji se talože u ćeliji ili van nje tokom amiloidoze, Alchajmerove bolesti itd. Ponekad prethodno strukturirani sličan molekul može poslužiti kao matriks, a pri tome U slučaju, ako se primarno strukturiranje dogodi pogrešno, svi naredni molekuli će također biti defektni. Ova situacija se javlja kod tzv. prionskih bolesti (scrapie kod ovaca, bijesnih krava, kuru, Creutzfeldt-Jakobova bolest kod ljudi), kada defekt jednog od membranskih proteina nervne ćelije uzrokuje naknadno nakupljanje inertnih masa unutar ćelije. i poremećaj njegovih vitalnih funkcija.
Do poremećaja procesa sinteze u ćeliji može doći u različitim fazama: transkripcija RNK u jezgru, translacija polipeptida u ribosomima, posttranslacijska modifikacija, hipermetilacija i glikozilacija bež molekula, transport i distribucija proteina u ćeliji i njihovo uklanjanje spolja. U ovom slučaju može se uočiti povećanje ili smanjenje broja ribozoma, raspad poliribosoma, širenje cisterni granularnog endoplazmatskog retikuluma, gubitak ribozoma i stvaranje vezikula i vakuola. Dakle, kada se otrova blijedim gnjurcem, enzim RNA polimeraza je oštećen, što remeti transkripciju. Toksin difterije, deaktivirajući faktor elongacije, remeti procese translacije, uzrokujući oštećenje miokarda. Uzrok poremećaja u sintezi nekih specifičnih proteinskih molekula mogu biti infektivni agensi. Na primjer, herpes virusi inhibiraju sintezu i ekspresiju molekula MHC antigena, što im omogućava da djelimično izbjegnu imunološku kontrolu; bacili kuge - sintezu medijatora akutne upale. Pojava neobičnih proteina može zaustaviti njihov daljnji razgradnju i dovesti do nakupljanja inertnog ili čak toksičnog materijala. Ovo se u određenoj mjeri može olakšati poremećajem procesa raspadanja.
Poremećaj procesa propadanja
Istovremeno sa sintezom proteina u ćeliji, kontinuirano dolazi do njegovog razlaganja. U normalnim uslovima, ovo ima važan regulatorni i formativni značaj, na primer, tokom aktivacije neaktivnih oblika enzima, proteinskih hormona i proteina mitotičkog ciklusa. Normalan rast i razvoj ćelije zahtevaju fino kontrolisanu ravnotežu između sinteze i razgradnje proteina i organela. Međutim, u procesu sinteze proteina, zbog grešaka u radu aparata za sintezu, abnormalnog strukturiranja proteinske molekule i njenog oštećenja kemijskim i bakterijskim agensima, konstantno nastaje prilično veliki broj neispravnih molekula. Prema nekim procjenama, njihov udio je oko trećine svih sintetiziranih proteina.
Ćelije sisara imaju nekoliko glavnih načini uništavanja proteina: kroz lizozomalne proteaze (pentidne hidrolaze), proteinaze zavisne od kalcijuma (endopeptidaze) i proteazomski sistem. Osim toga, postoje i specijalizirane proteinaze, kao što su kaspaze. Glavna organela u kojoj dolazi do razgradnje tvari u eukariotskim stanicama je lizozom, koji sadrži brojne hidrolitičke enzime. Zbog procesa endocitoze i različitih tipova autofagije u lizosomima i fagolizosomima uništavaju se kako defektni proteinski molekuli tako i cijele organele: oštećeni mitohondriji, dijelovi plazma membrane, neki ekstracelularni proteini i sadržaj sekretornih granula.
Važan mehanizam za razgradnju proteina je proteasom, multikatalitička proteinazna struktura složene strukture lokalizovana u citosolu, jezgru, endoplazmatskom retikulumu i na ćelijskoj membrani. Ovaj enzimski sistem je odgovoran za razgradnju oštećenih proteina, kao i zdravih proteina koji se moraju ukloniti za normalnu funkciju ćelija. U ovom slučaju, proteini koji se uništavaju se preliminarno kombinuju sa specifičnim polipeptidom, ubikvitinom. Međutim, ne-ubikvitinirani proteini također mogu biti djelomično uništeni u proteazomima. Razgradnja proteinskih molekula u proteazomima u kratke polipeptide (obrada) sa njihovom naknadnom prezentacijom zajedno sa molekulima tipa I MHC važna je karika u imunološkoj kontroli antigenske homeostaze u tijelu. Kada je funkcija proteasoma oslabljena, akumuliraju se oštećeni i nepotrebni proteini, što prati starenje stanica. Kršenje razgradnje proteina ovisnih o ciklinu dovodi do poremećaja diobe stanica, degradacije sekretornih proteina - do razvoja cistofibroze. Nasuprot tome, povećanje funkcije proteasoma prati iscrpljivanje organizma (AIDS, rak).
Uz genetski uslovljene poremećaje razgradnje proteina, organizam nije održiv i umire u ranim fazama embriogeneze. Ako je razgradnja masti ili ugljikohidrata poremećena, dolazi do bolesti skladištenja (tezaurizma). U tom slučaju se unutar ćelije nakuplja prekomjerna količina određenih tvari ili produkata njihovog nepotpunog razlaganja – lipida, polisaharida, što značajno narušava funkciju stanice. Najčešće se to opaža u epitelnim ćelijama jetre (hepatocitima), neuronima, fibroblastima i makrofagocitima.
Stečeni poremećaji procesa razgradnje tvari mogu nastati kao posljedica patoloških procesa (na primjer, proteina, masti, ugljikohidrata i pigmentne degeneracije) i praćeni su stvaranjem neobičnih tvari. Poremećaji u sistemu lizozomske proteolize dovode do smanjene adaptacije tokom gladovanja ili pojačanog stresa, te do pojave određenih endokrinih disfunkcija – sniženog nivoa insulina, tireoglobulina, citokina i njihovih receptora. Poremećaj razgradnje proteina usporava brzinu zacjeljivanja rana, uzrokuje razvoj ateroskleroze i utječe na imunološki odgovor. S hipoksijom, promjenama unutarćelijskog pH, ozljedama zračenja, koje karakterizira povećana peroksidacija membranskih lipida, kao i pod utjecajem lizosomotropnih tvari - bakterijskih endotoksina, metabolita toksičnih gljivica (sporofusarin), kristala silicijum oksida - stabilnost membrane lizosoma promjenama, aktivirani lizosomski enzimi se oslobađaju u citoplazmu, što uzrokuje uništavanje staničnih struktura i njihovu smrt.
Veza organizma sa okolinom, sa fizičko-hemijske tačke gledišta, je otvoren sistem, odnosno sistem u kome se odvijaju biohemijski procesi. Polazne supstance dolaze iz okoline, a supstance koje se takođe kontinuirano formiraju prenose se van. Ravnoteža između brzine i koncentracije produkata višesmjernih reakcija u tijelu je uvjetna, imaginarna, budući da se unos i uklanjanje tvari ne zaustavlja. Kontinuirana povezanost sa okolinom omogućava nam da posmatramo živi organizam kao otvoreni sistem.
Za sve žive ćelije izvor energije je Sunce. Biljne ćelije hvataju energiju sunčeve svetlosti uz pomoć hlorofila, koristeći je za reakcije asimilacije tokom procesa fotosinteze. Ćelije životinja, gljiva i bakterija koriste sunčevu energiju indirektno, prilikom razgradnje organskih tvari koje sintetiziraju zemaljske biljke.
Neki od hranljivih sastojaka ćelije se razgrađuju tokom ćelijskog disanja, čime se snabdeva energijom neophodnom za različite vrste ćelijske aktivnosti. Ovaj proces se odvija u organelama zvanim mitohondrije. Mitohondrije se sastoje od dvije membrane: vanjske, koja odvaja organelu od citoplazme, i unutrašnje, koja formira brojne nabore. Glavni proizvod disanja je ATP. Napušta mitohondrije i koristi se kao izvor energije za mnoge hemijske reakcije u citoplazmi i ćelijskoj membrani. Ako je kisik potreban za stanično disanje, tada se disanje naziva aerobno, ali ako se reakcije odvijaju u nedostatku kisika, onda govorimo o anaerobnom disanju.
Za bilo koju vrstu rada koji se obavlja u ćeliji, energija se koristi u jednom jedinom obliku - u obliku energije iz fosfatnih veza ATP-a. ATP je lako pokretno jedinjenje. Formiranje ATP-a događa se na unutrašnjoj membrani mitohondrija. ATP se sintetiše u svim ćelijama tokom disanja zahvaljujući energiji oksidacije ugljenih hidrata, masti i drugih organskih materija. U zelenim biljnim ćelijama, glavna količina ATP-a se sintetizira u hloroplastima zahvaljujući sunčevoj energiji. Tokom fotosinteze proizvode mnogo puta više ATP-a nego mitohondrije. ATP se razgrađuje kidanjem veza fosfor-kiseonik i oslobađanjem energije. To se događa pod djelovanjem enzima ATPaze tokom hidrolize ATP-a - dodavanjem vode uz eliminaciju molekula fosforne kiseline. Kao rezultat, ATP se pretvara u ADP, a ako se odvoje dva molekula fosforne kiseline, onda u AMP. Reakcija eliminacije svakog gram-molekula kiseline je praćena oslobađanjem 40 kJ. Ovo je vrlo velika izlazna energija, zbog čega se veze fosfora i kisika ATP-a obično nazivaju makroergističkim (visokoenergetskim).
Upotreba ATP-a u reakcijama plastične izmjene provodi se njihovim spajanjem s hidrolizom ATP-a. Molekuli različitih supstanci se nabijaju energijom vezivanjem fosforne grupe koja se oslobađa tokom hidrolize iz molekula ATP-a, odnosno fosforilacijom.
Posebnost derivata fosfata je u tome što ne mogu napustiti ćeliju, iako njihovi "ispušteni" oblici slobodno prolaze kroz membranu. Zahvaljujući tome, fosforilirani molekuli ostaju u ćeliji sve dok se ne iskoriste u odgovarajućim reakcijama.
Obrnuti proces pretvaranja ADP u ATP događa se dodavanjem molekula fosforne kiseline ADP-u, oslobađanjem vode i apsorbiranjem velike količine energije.
Dakle, ATP je univerzalni i direktni izvor energije za ćelijsku aktivnost. Ovo stvara jedinstveni ćelijski bazen energije i omogućava njenu redistribuciju i transport iz jednog područja ćelije u drugo.
Prijenos fosfatne grupe igra važnu ulogu u kemijskim reakcijama kao što je sastavljanje makromolekula iz monomera. Na primjer, aminokiseline se mogu kombinovati u peptide tek nakon što su prethodno fosforilirane. Mehanički procesi kontrakcije ili kretanja, transport otopljene supstance protiv gradijenta koncentracije i drugi procesi uključuju potrošnju energije pohranjene u ATP-u.
Proces energetskog metabolizma može se predstaviti na sljedeći način. Visokomolekularne organske tvari u citoplazmi se enzimski, hidrolizom, pretvaraju u jednostavnije od kojih se sastoje: proteini - u aminokiseline, poli- i disaharidi - u monosaharide (+ glukoza), masti u glicerol i masne kiseline. Nema oksidativnih procesa, oslobađa se malo energije koja se ne koristi i prelazi u termalni oblik. Većina ćelija prvo koristi ugljikohidrate. Polisaharidi (škrob u biljkama i glikogen kod životinja) hidroliziraju se u glukozu. Oksidacija glukoze se odvija u tri faze: glikoliza, oksidativna dekarboksilacija (Krebsov ciklus - ciklus limunske kiseline) i oksidativna fosforilacija (respiratorni lanac). U citoplazmi se javlja glikoliza, zbog koje se jedan molekul glukoze dijeli na dva molekula pirogrožđane kiseline uz oslobađanje dva molekula ATP-a. U nedostatku kisika, pirogrožđana kiselina se pretvara u etanol (fermentacija) ili mliječnu kiselinu (anaerobno disanje).
Kada se glikoliza dogodi u životinjskim stanicama, molekula glukoze sa šest ugljika razlaže se na dva molekula mliječne kiseline. Ovaj proces je višestepeni. Izvodi ga uzastopno 13 enzima. Tokom alkoholne fermentacije iz molekula glukoze nastaju dva molekula etanola i dva molekula CO2.
Glikoliza je faza zajednička za anaerobno i aerobno disanje; druge dvije se javljaju samo u aerobnim uvjetima. Proces oksidacije bez kisika, u kojem se oslobađa i koristi samo dio energije metabolita, konačan je za anaerobne organizme. U prisustvu kisika, pirogrožđana kiselina prelazi u mitohondrije, gdje se kao rezultat niza uzastopnih reakcija potpuno aerobno oksidira u H2O i CO2 uz istovremenu fosforilaciju ADP-a u ATP. U ovom slučaju, dva ATP molekula se proizvode glikolizom, dva Krebsovim ciklusom, a 34 respiratornim lancem. Neto prinos za potpunu oksidaciju jednog molekula glukoze u H2O i CO2 je 38 molekula.
Tako se kod aerobnih organizama konačna razgradnja organskih tvari vrši oksidacijom atmosferskim kisikom do jednostavnih anorganskih tvari: CO2 i H2O. Ovaj proces se odvija na kristama mitohondrija. U tom slučaju se oslobađa maksimalna količina slobodne energije, čiji je značajan dio rezervisan u molekulima ATP-a. Lako je vidjeti da aerobna oksidacija u najvećoj mjeri daje ćeliji besplatnu energiju.
Kao rezultat katabolizma, energetski bogati ATP molekuli se akumuliraju u ćeliji, a CO2 i višak vode oslobađaju se u vanjsko okruženje.
Molekuli šećera koji nisu potrebni za disanje mogu se pohraniti u ćeliji. Višak lipida se ili razgrađuje, nakon čega proizvodi njihovog razgradnje ulaze u mitohondrije kao supstrat za disanje, ili se talože kao rezerve u citoplazmi u obliku masnih kapljica. Proteini se grade od aminokiselina koje ulaze u ćeliju. Sinteza proteina se odvija u organelama zvanim ribosomi. Svaki ribosom se sastoji od dvije podčestice - velike i male: obje podčestice uključuju proteinske molekule i molekule RNK.
Ribozomi su često vezani za poseban membranski sistem koji se sastoji od cisterni i vezikula - takozvani endoplazmatski retikulum (ER); u ćelijama koje proizvode mnogo proteina, endoplazmatski retikulum je često vrlo dobro razvijen i prekriven ribosomima. Neki enzimi su efikasni samo ako su vezani za membranu. Većina enzima uključenih u sintezu lipida nalazi se ovdje. Dakle, endoplazmatski retikulum je kao neka vrsta ćelijskog radnog stola.
Osim toga, ER dijeli citoplazmu u zasebne odjeljke, tj. odvaja različite kemijske procese koji se istovremeno odvijaju u citoplazmi, i na taj način smanjuje vjerovatnoću da će ti procesi interferirati jedan s drugim.
Proizvodi koje proizvodi određena ćelija često se koriste izvan ćelije. U takvim slučajevima, proteini sintetizirani na ribosomima prolaze kroz membrane endoplazmatskog retikuluma i pakuju se u membranske vezikule koje se formiraju oko njih, koje se zatim odvajaju od ER. Ove vezikule, spljoštene i naslagane jedna na drugu, poput naslaganih palačinki, formiraju karakterističnu strukturu koja se naziva Golgijev kompleks ili Golgijev aparat. Tokom boravka u Golgijevom aparatu, proteini prolaze kroz određene promjene. Kada dođe vrijeme da napuste ćeliju, membranski mjehurići se spajaju sa ćelijskom membranom i prazne se, izlivajući svoj sadržaj, tj. do izlučivanja dolazi egzocitozom.
Golgijev aparat također proizvodi lizozome - membranske vrećice koje sadrže probavne enzime. Otkrivanje kako ćelija proizvodi, pakuje i izvozi određene proteine i kako "zna" koje proteine treba da zadrži za sebe, jedna je od najfascinantnijih grana moderne citologije.
Membrane bilo koje ćelije se stalno kreću i mijenjaju. ER membrane se kreću polako kroz ćeliju. Pojedinačni dijelovi ovih membrana se odvajaju i formiraju vezikule, koje privremeno postaju dio Golgijevog aparata, a zatim se procesom egzocitoze spajaju sa ćelijskom membranom.
Kasnije se membranski materijal vraća u citoplazmu, gdje se ponovo koristi.