 Eingang
EingangInteraktion der Zelle mit der Umgebung. Beziehung zwischen Organismus und Umwelt. Entwicklung von Keimzellen bei Pflanzen und Tieren
Wir laden Sie ein, sich mit den Materialien und vertraut zu machen.
: Zellulosemembran, Membran, Zytoplasma mit Organellen, Zellkern, Vakuolen mit Zellsaft.Das Vorhandensein von Plastiden ist das Hauptmerkmal einer Pflanzenzelle.
Funktionen der Zellmembran- bestimmt die Form der Zelle, schützt vor Umwelteinflüssen.
Plasma Membran- Ein dünner Film, bestehend aus interagierenden Lipid- und Proteinmolekülen, grenzt den inneren Inhalt von der äußeren Umgebung ab, sorgt durch Osmose und aktiven Transport für den Transport von Wasser, Mineralien und organischen Substanzen in die Zelle und entfernt außerdem Abfallprodukte.
Zytoplasma- Die innere halbflüssige Umgebung der Zelle, in der sich der Zellkern und die Organellen befinden, sorgt für Verbindungen zwischen ihnen und ist an grundlegenden Lebensprozessen beteiligt.
Endoplasmatisches Retikulum- ein Netzwerk verzweigter Kanäle im Zytoplasma. Es ist an der Synthese von Proteinen, Lipiden und Kohlenhydraten sowie am Stofftransport beteiligt. Ribosomen sind im ER oder im Zytoplasma befindliche Körper, die aus RNA und Protein bestehen und an der Proteinsynthese beteiligt sind. EPS und Ribosomen sind ein einziger Apparat für die Synthese und den Transport von Proteinen.
Mitochondrien- Organellen, die durch zwei Membranen vom Zytoplasma abgegrenzt sind. In ihnen werden organische Substanzen oxidiert und ATP-Moleküle unter Beteiligung von Enzymen synthetisiert. Vergrößerung der Oberfläche der inneren Membran, auf der sich Enzyme befinden, durch Cristae. ATP ist eine energiereiche organische Substanz.
Plastiden(Chloroplasten, Leukoplasten, Chromoplasten), ihr Inhalt in der Zelle ist das Hauptmerkmal des Pflanzenorganismus. Chloroplasten sind Plastiden, die den grünen Farbstoff Chlorophyll enthalten, der Lichtenergie absorbiert und damit organische Substanzen aus Kohlendioxid und Wasser synthetisiert. Chloroplasten sind vom Zytoplasma durch zwei Membranen getrennt, zahlreiche Auswüchse – Grana auf der inneren Membran, in der sich Chlorophyllmoleküle und Enzyme befinden.
Golgi-Komplex- ein System von Hohlräumen, die durch eine Membran vom Zytoplasma abgegrenzt sind. Die Ansammlung von Proteinen, Fetten und Kohlenhydraten darin. Durchführung der Synthese von Fetten und Kohlenhydraten auf Membranen.
Lysosomen- Körper, die durch eine einzige Membran vom Zytoplasma abgegrenzt sind. Die darin enthaltenen Enzyme beschleunigen den Abbau komplexer Moleküle in einfache: Proteine in Aminosäuren, komplexe Kohlenhydrate in einfache, Lipide in Glycerin und Fettsäuren und zerstören auch abgestorbene Zellteile und ganze Zellen.
Vakuolen- mit Zellsaft gefüllte Hohlräume im Zytoplasma, ein Ort der Ansammlung von Reservenährstoffen und Schadstoffen; Sie regulieren den Wassergehalt in der Zelle.
Kern- der Hauptteil der Zelle, außen mit einer zweimembranigen, porendurchbohrten Kernhülle bedeckt. Stoffe gelangen in den Kern und werden durch die Poren aus diesem entfernt. Chromosomen sind Träger erblicher Informationen über die Eigenschaften eines Organismus, die Hauptstrukturen des Zellkerns, die jeweils aus einem DNA-Molekül kombiniert mit Proteinen bestehen. Der Zellkern ist der Ort der DNA-, mRNA- und rRNA-Synthese.

Das Vorhandensein einer äußeren Membran, eines Zytoplasmas mit Organellen und eines Zellkerns mit Chromosomen.
Äußere oder Plasmamembran- grenzt den Zellinhalt von der Umgebung (andere Zellen, Interzellularsubstanz) ab, besteht aus Lipid- und Proteinmolekülen, sorgt für die Kommunikation zwischen Zellen, den Stofftransport in die Zelle (Pinozytose, Phagozytose) und aus der Zelle heraus.
Zytoplasma- die innere halbflüssige Umgebung der Zelle, die für die Kommunikation zwischen dem Zellkern und den darin befindlichen Organellen sorgt. Die wichtigsten Lebensprozesse finden im Zytoplasma statt.
Zellorganellen:
1) endoplasmatisches Retikulum (ER)- ein System verzweigter Tubuli, beteiligt sich an der Synthese von Proteinen, Lipiden und Kohlenhydraten sowie am Stofftransport in der Zelle;
2) Ribosomen- rRNA-haltige Körper befinden sich im ER und im Zytoplasma und sind an der Proteinsynthese beteiligt. EPS und Ribosomen sind ein einziger Apparat für die Proteinsynthese und den Proteintransport;
3) Mitochondrien- „Kraftwerke“ der Zelle, vom Zytoplasma durch zwei Membranen abgegrenzt. Das Innere bildet Cristae (Falten), wodurch seine Oberfläche vergrößert wird. Enzyme auf den Cristae beschleunigen die Oxidation organischer Substanzen und die Synthese energiereicher ATP-Moleküle;
4) Golgi-Komplex- eine Gruppe von Hohlräumen, die durch eine Membran vom Zytoplasma abgegrenzt sind und mit Proteinen, Fetten und Kohlenhydraten gefüllt sind, die entweder in lebenswichtigen Prozessen verwendet oder aus der Zelle entfernt werden. Die Membranen des Komplexes übernehmen die Synthese von Fetten und Kohlenhydraten;
5) Lysosomen- Mit Enzymen gefüllte Körper beschleunigen den Abbau von Proteinen in Aminosäuren, Lipiden in Glycerin und Fettsäuren, Polysacchariden in Monosaccharide. In Lysosomen werden abgestorbene Zellteile, also ganze Zellen, zerstört.
Zelluläre Einschlüsse- Ansammlung von Reservenährstoffen: Proteine, Fette und Kohlenhydrate.
Kern- der wichtigste Teil der Zelle. Es ist mit einer Doppelmembranhülle mit Poren bedeckt, durch die einige Substanzen in den Zellkern und andere in das Zytoplasma eindringen. Chromosomen sind die Hauptstrukturen des Zellkerns und Träger erblicher Informationen über die Eigenschaften des Organismus. Es wird bei der Teilung der Mutterzelle auf Tochterzellen und von Keimzellen auf Tochterorganismen übertragen. Der Zellkern ist der Ort der DNA-, mRNA- und rRNA-Synthese.
Übung:
Erklären Sie, warum Organellen als spezialisierte Zellstrukturen bezeichnet werden.
Antwort: Organellen werden als spezialisierte Zellstrukturen bezeichnet, da sie streng definierte Funktionen erfüllen, Erbinformationen im Zellkern gespeichert werden, ATP in Mitochondrien synthetisiert wird, Photosynthese in Chloroplasten stattfindet usw.
Wenn Sie Fragen zur Zytologie haben, können Sie sich an uns wenden
Theorie zu Aufgabe 5 des Einheitlichen Staatsexamens in Biologie
Zellstruktur. Die Beziehung zwischen der Struktur und den Funktionen der Teile und Organellen einer Zelle ist die Grundlage ihrer Integrität
Zellstruktur
Struktur prokaryotischer und eukaryotischer Zellen
Die wichtigsten Strukturbestandteile von Zellen sind die Plasmamembran, das Zytoplasma und der Erbapparat. Abhängig von den Merkmalen der Organisation werden zwei Haupttypen von Zellen unterschieden: prokaryotische und eukaryotische. Der Hauptunterschied zwischen prokaryotischen Zellen und eukaryotischen Zellen besteht in der Organisation ihres Erbapparates: Bei Prokaryoten befindet er sich direkt im Zytoplasma (dieser Bereich des Zytoplasmas wird als „Zytoplasma“ bezeichnet). Nukleoid) und ist nicht durch Membranstrukturen von ihm getrennt, während bei Eukaryoten der Großteil der DNA im Zellkern konzentriert ist, der von einer Doppelmembran umgeben ist. Darüber hinaus ist die genetische Information prokaryotischer Zellen, die sich im Nukleoid befindet, in einem zirkulären DNA-Molekül geschrieben, während bei Eukaryoten die DNA-Moleküle offen sind.
Im Gegensatz zu Eukaryoten enthält das Zytoplasma prokaryotischer Zellen auch eine geringe Anzahl von Organellen, während eukaryotische Zellen durch eine erhebliche Vielfalt dieser Strukturen gekennzeichnet sind.
Struktur und Funktionen biologischer Membranen
Die Struktur der Biomembran. Die zellbegrenzenden Membranen und Membranorganellen eukaryontischer Zellen haben eine gemeinsame chemische Zusammensetzung und Struktur. Dazu gehören Lipide, Proteine und Kohlenhydrate. Membranlipide werden hauptsächlich durch Phospholipide und Cholesterin repräsentiert. Die meisten Membranproteine sind komplexe Proteine, beispielsweise Glykoproteine. Kohlenhydrate kommen in der Membran nicht unabhängig vor, sondern sind mit Proteinen und Lipiden verbunden. Die Dicke der Membranen beträgt 7–10 nm.
Nach dem derzeit allgemein anerkannten Fluidmosaikmodell der Membranstruktur bilden Lipide eine Doppelschicht, bzw lipiddoppelschicht, bei dem die hydrophilen „Köpfe“ der Lipidmoleküle nach außen zeigen und die hydrophoben „Schwänze“ in der Membran verborgen sind. Diese „Schwänze“ sorgen aufgrund ihrer Hydrophobie für die Trennung der wässrigen Phasen der inneren Umgebung der Zelle und ihrer Umgebung. Proteine sind durch verschiedene Arten von Wechselwirkungen mit Lipiden verbunden. Einige Proteine befinden sich auf der Oberfläche der Membran. Solche Proteine heißen peripher, oder oberflächlich. Andere Proteine sind teilweise oder vollständig in die Membran eingetaucht – das sind Integral, oder untergetauchte Proteine. Membranproteine erfüllen Struktur-, Transport-, Katalyse-, Rezeptor- und andere Funktionen.
Membranen sind nicht wie Kristalle; ihre Bestandteile sind ständig in Bewegung, wodurch Lücken zwischen Lipidmolekülen entstehen – Poren, durch die verschiedene Substanzen in die Zelle eindringen oder diese verlassen können.
Biologische Membranen unterscheiden sich in ihrer Lage in der Zelle, ihrer chemischen Zusammensetzung und ihren Funktionen. Die wichtigsten Membrantypen sind Plasma- und interne Membranen. Plasma Membran enthält etwa 45 % Lipide (einschließlich Glykolipide), 50 % Proteine und 5 % Kohlenhydrate. Über die Oberfläche der Membran ragen Kohlenhydratketten hinaus, die Teil komplexer Proteine-Glykoproteine und komplexer Lipide-Glykolipide sind. Plasmalemma-Glykoproteine sind äußerst spezifisch. Sie dienen beispielsweise der gegenseitigen Erkennung von Zellen, einschließlich Spermium und Eizelle.
Auf der Oberfläche tierischer Zellen bilden Kohlenhydratketten eine dünne Oberflächenschicht – Glykokalyx. Es wird in fast allen tierischen Zellen nachgewiesen, sein Expressionsgrad variiert jedoch (10–50 µm). Die Glykokalyx sorgt für die direkte Kommunikation zwischen der Zelle und der äußeren Umgebung, wo die extrazelluläre Verdauung stattfindet; Rezeptoren befinden sich in der Glykokalyx. Neben dem Plasmalemma sind auch die Zellen von Bakterien, Pflanzen und Pilzen von Zellmembranen umgeben.
Interne Membranen Eukaryontische Zellen grenzen verschiedene Teile der Zelle ab und bilden eigenartige „Kompartimente“ – Fächer, was die Trennung verschiedener Stoffwechsel- und Energieprozesse fördert. Sie können sich in ihrer chemischen Zusammensetzung und ihren Funktionen unterscheiden, ihr allgemeiner Strukturplan bleibt jedoch derselbe.
Membranfunktionen:
- Begrenzend. Die Idee dahinter ist, dass sie den Innenraum der Zelle von der Außenumgebung trennen. Die Membran ist semipermeabel, das heißt, nur die Stoffe, die die Zelle benötigt, können sie ungehindert passieren, und es gibt Mechanismen zum Transport der notwendigen Stoffe.
- Rezeptor. Es ist in erster Linie mit der Wahrnehmung von Umweltsignalen und der Übertragung dieser Informationen in die Zelle verbunden. Für diese Funktion sind spezielle Rezeptorproteine verantwortlich. Membranproteine sind auch für die zelluläre Erkennung nach dem „Freund-Feind“-Prinzip sowie für die Bildung interzellulärer Verbindungen verantwortlich, von denen die Synapsen von Nervenzellen am besten untersucht sind.
- Katalytisch. Auf den Membranen befinden sich zahlreiche Enzymkomplexe, wodurch auf ihnen intensive Syntheseprozesse ablaufen.
- Energieumwandlung. Verbunden mit der Bildung von Energie, ihrer Speicherung in Form von ATP und ihrem Verbrauch.
- Kompartimentierung. Membranen begrenzen außerdem den Raum innerhalb der Zelle und trennen so die Ausgangsstoffe der Reaktion und die Enzyme, die die entsprechenden Reaktionen durchführen können.
- Bildung interzellulärer Kontakte. Obwohl die Dicke der Membran so gering ist, dass sie mit bloßem Auge nicht erkennbar ist, dient sie einerseits als recht zuverlässige Barriere für Ionen und Moleküle, insbesondere wasserlösliche, und andererseits , sorgt für deren Transport in die Zelle hinein und aus ihr heraus.
- Transport.

Membrantransport. Da Zellen als elementare biologische Systeme offene Systeme sind, ist zur Sicherstellung von Stoffwechsel und Energie, zur Aufrechterhaltung der Homöostase, des Wachstums, der Reizbarkeit und anderer Prozesse die Übertragung von Stoffen durch die Membran – der Membrantransport – erforderlich. Derzeit wird der Stofftransport durch die Zellmembran in aktive, passive, Endo- und Exozytose unterteilt.
Passiver Transport- Hierbei handelt es sich um eine Transportart, die ohne Energieaufwand von höheren zu niedrigeren Konzentrationen erfolgt. Lipidlösliche kleine unpolare Moleküle (O 2, CO 2) dringen leicht in die Zelle ein einfache Diffusion. In Lipiden unlösliche Stoffe, einschließlich geladener kleiner Partikel, werden von Trägerproteinen aufgenommen oder passieren spezielle Kanäle (Glukose, Aminosäuren, K +, PO 4 3-). Diese Art des passiven Transports wird aufgerufen erleichterte Diffusion. Wasser gelangt durch Poren in der Lipidphase sowie durch spezielle, mit Proteinen ausgekleidete Kanäle in die Zelle. Den Transport von Wasser durch eine Membran nennt man durch Osmose.

Osmose ist im Leben einer Zelle äußerst wichtig, denn wenn sie in eine Lösung mit einer höheren Salzkonzentration als in der Zelllösung gegeben wird, beginnt Wasser aus der Zelle auszutreten und das Volumen des lebenden Inhalts beginnt abzunehmen. Bei tierischen Zellen schrumpft die Zelle als Ganzes, und bei pflanzlichen Zellen bleibt das Zytoplasma hinter der Zellwand zurück, was als „Zellzellwand“ bezeichnet wird Plasmolyse. Wenn eine Zelle in eine Lösung gegeben wird, die weniger konzentriert ist als das Zytoplasma, erfolgt der Wassertransport in die entgegengesetzte Richtung – in die Zelle hinein. Der Dehnbarkeit der Zytoplasmamembran sind jedoch Grenzen gesetzt, und eine tierische Zelle reißt irgendwann, während eine Pflanzenzelle dies aufgrund ihrer starken Zellwand nicht zulässt. Als Phänomen bezeichnet man das Füllen des gesamten Innenraums einer Zelle mit Zellinhalt Deplasmolyse. Bei der Zubereitung von Medikamenten, insbesondere bei intravenöser Verabreichung, sollte die intrazelluläre Salzkonzentration berücksichtigt werden, da diese zu einer Schädigung der Blutzellen führen kann (hierfür wird Kochsalzlösung mit einer Konzentration von 0,9 % Natriumchlorid verwendet). Dies ist nicht weniger wichtig bei der Kultivierung von Zellen und Geweben sowie tierischen und pflanzlichen Organen.

Aktiven Transport geht mit dem Verbrauch von ATP-Energie von einer niedrigeren Konzentration einer Substanz zu einer höheren über. Dies geschieht mithilfe spezieller Pumpproteine. Proteine pumpen K + , Na + , Ca 2+ und andere Ionen durch die Membran, was den Transport essentieller organischer Substanzen sowie die Entstehung von Nervenimpulsen usw. fördert.
Endozytose- Dies ist ein aktiver Prozess der Aufnahme von Substanzen durch die Zelle, bei dem die Membran Einstülpungen und dann Membranvesikel bildet - Phagosomen, die die absorbierten Objekte enthalten. Dann verschmilzt das primäre Lysosom mit dem Phagosom und bildet sich sekundäres Lysosom, oder Phagolysosom, oder Verdauungsvakuole. Der Inhalt der Vesikel wird durch Lysosomenenzyme verdaut und die Abbauprodukte werden von der Zelle absorbiert und assimiliert. Unverdaute Rückstände werden durch Exozytose aus der Zelle entfernt. Es gibt zwei Haupttypen der Endozytose: Phagozytose und Pinozytose.
Phagozytose ist der Prozess des Einfangens fester Partikel durch die Zelloberfläche und der Absorption durch die Zelle Pinozytose- Flüssigkeiten. Phagozytose kommt vor allem in tierischen Zellen (einzellige Tiere, menschliche Leukozyten) vor, sie liefert deren Nahrung und schützt oft den Körper. Durch Pinozytose werden Proteine, Antigen-Antikörper-Komplexe bei Immunreaktionen etc. aufgenommen. Viele Viren gelangen jedoch auch durch Pinozytose oder Phagozytose in die Zelle. In Pflanzen- und Pilzzellen ist eine Phagozytose praktisch unmöglich, da sie von haltbaren Zellmembranen umgeben sind.

Exozytose- ein zur Endozytose umgekehrter Prozess. Auf diese Weise werden unverdaute Nahrungsreste aus den Verdauungsvakuolen gelöst und für das Leben der Zelle und des gesamten Körpers notwendige Stoffe entfernt. Beispielsweise erfolgt die Übertragung von Nervenimpulsen durch die Freisetzung chemischer Botenstoffe durch das Neuron, das den Impuls sendet – Vermittler, und in Pflanzenzellen werden auf diese Weise Hilfskohlenhydrate der Zellmembran ausgeschieden.
Zellwände von Pflanzenzellen, Pilzen und Bakterien. Außerhalb der Membran kann die Zelle ein starkes Gerüst absondern – Zellmembran, oder Zellenwand.
Bei Pflanzen ist die Basis die Zellwand Zellulose, verpackt in Bündeln von 50-100 Molekülen. Die Zwischenräume sind mit Wasser und anderen Kohlenhydraten gefüllt. Die pflanzliche Zellwand ist von Tubuli durchzogen - Plasmodesmen, durch die die Membranen des endoplasmatischen Retikulums verlaufen. Plasmodesmen übernehmen den Stofftransport zwischen Zellen. Der Transport von Stoffen, beispielsweise Wasser, kann jedoch auch entlang der Zellwände selbst erfolgen. Im Laufe der Zeit reichern sich in der Zellwand von Pflanzen verschiedene Stoffe, darunter Tannine oder fettähnliche Stoffe, an, was zur Verholzung bzw. Suberisierung der Zellwand selbst, zur Wasserverdrängung und zum Absterben des Zellinhalts führt. Zwischen den Zellwänden benachbarter Pflanzenzellen befinden sich geleeartige Abstandshalter – Mittelplatten, die sie zusammenhalten und den Pflanzenkörper als Ganzes zementieren. Sie werden erst während der Fruchtreife und beim Laubfall zerstört.

Die Zellwände von Pilzzellen werden gebildet Chitin- ein stickstoffhaltiges Kohlenhydrat. Sie sind ziemlich stark und bilden das äußere Skelett der Zelle, verhindern aber dennoch, wie bei Pflanzen, die Phagozytose.
Bei Bakterien enthält die Zellwand Kohlenhydrate mit Peptidfragmenten – Murein Allerdings variiert sein Gehalt je nach Bakteriengruppe erheblich. Auch andere Polysaccharide können auf der Zellwand abgesondert werden und dort eine Schleimkapsel bilden, die Bakterien vor äußeren Einflüssen schützt.
Die Membran bestimmt die Form der Zelle, dient als mechanische Stütze, übt eine Schutzfunktion aus, sorgt für die osmotischen Eigenschaften der Zelle, begrenzt die Dehnung des lebenden Inhalts und verhindert das Aufbrechen der Zelle, das durch das Eindringen von Wasser zunimmt . Darüber hinaus überwinden Wasser und darin gelöste Stoffe die Zellwand, bevor sie in das Zytoplasma gelangen oder umgekehrt, wenn sie es verlassen, während Wasser schneller durch die Zellwände transportiert wird als durch das Zytoplasma.
Zytoplasma
Zytoplasma- Dies ist der interne Inhalt der Zelle. Alle Zellorganellen, der Zellkern und verschiedene Abfallprodukte sind darin eingetaucht.
Das Zytoplasma verbindet alle Teile der Zelle miteinander und in ihm finden zahlreiche Stoffwechselreaktionen statt. Das Zytoplasma ist von der Umgebung getrennt und durch Membranen in Kompartimente unterteilt, d. h. Zellen haben eine Membranstruktur. Es kann in zwei Zuständen vorliegen – Sol und Gel. Sol- Dies ist ein halbflüssiger, geleeartiger Zustand des Zytoplasmas, in dem lebenswichtige Prozesse am intensivsten ablaufen, und Gel- ein dichterer, gallertartiger Zustand, der das Auftreten chemischer Reaktionen und den Stofftransport behindert.
Der flüssige Teil des Zytoplasmas ohne Organellen wird genannt Hyaloplasma. Hyaloplasma oder Zytosol ist eine kolloidale Lösung, in der sich eine Art Suspension aus relativ großen Partikeln, zum Beispiel Proteinen, befindet, die von Dipolen aus Wassermolekülen umgeben sind. Aufgrund der gleichen Ladung und der gegenseitigen Abstoßung kommt es zu keiner Ausfällung dieser Suspension.
Organoide
Organoide- Dies sind dauerhafte Bestandteile der Zelle, die bestimmte Funktionen erfüllen.
Abhängig von ihren strukturellen Merkmalen werden sie in Membran- und Nichtmembranmembranen unterteilt. Membran Organellen wiederum werden in Einzelmembranorganellen (endoplasmatisches Retikulum, Golgi-Komplex und Lysosomen) oder Doppelmembranorganellen (Mitochondrien, Plastiden und Zellkern) eingeteilt. Nicht-Membran Die Organellen sind Ribosomen, Mikrotubuli, Mikrofilamente und das Zellzentrum. Von den aufgeführten Organellen sind Prokaryoten nur Ribosomen inhärent.
Struktur und Funktionen des Kerns. Kern- ein großes Doppelmembranorganell, das in der Mitte der Zelle oder an ihrer Peripherie liegt. Die Abmessungen des Kerns können zwischen 3 und 35 Mikrometern liegen. Die Form des Kerns ist meist kugelförmig oder ellipsoid, es gibt aber auch stäbchenförmige, spindelförmige, bohnenförmige, gelappte und sogar segmentierte Kerne. Einige Forscher glauben, dass die Form des Zellkerns der Form der Zelle selbst entspricht.
Die meisten Zellen haben einen Kern, aber zum Beispiel können es in den Zellen der Leber und des Herzens zwei sein, und in mehreren Neuronen – bis zu 15. Skelettmuskelfasern enthalten normalerweise viele Kerne, aber sie sind keine Zellen im wahrsten Sinne des Wortes, da sie durch die Verschmelzung mehrerer Zellen entstehen.
Der Kern ist umgeben Atomhülle, und sein Innenraum ist gefüllt nuklearer Saft, oder Nukleoplasma (Karyoplasma), in dem sie eingetaucht sind Chromatin Und Nukleolus. Der Zellkern erfüllt so wichtige Funktionen wie die Speicherung und Übertragung von Erbinformationen sowie die Steuerung des Zelllebens.

Die Rolle des Zellkerns bei der Übertragung von Erbinformationen wurde in Experimenten mit der Grünalge Acetabularia überzeugend nachgewiesen. In einer einzelnen Riesenzelle, die eine Länge von 5 cm erreicht, werden eine Kappe, ein Stiel und ein Rhizoid unterschieden. Darüber hinaus enthält es nur einen Kern, der sich im Rhizoid befindet. In den 1930er Jahren transplantierte I. Hemmerling den Kern einer grün gefärbten Acetabularia-Art in das Rhizoid einer anderen braun gefärbten Art, aus dem der Kern entfernt worden war. Nach einiger Zeit wuchs an der Pflanze mit dem transplantierten Kern eine neue Kappe, wie bei den Kernspenderalgen. Gleichzeitig starb der Hut oder Stiel, der vom Rhizoid getrennt war und keinen Kern enthielt, nach einiger Zeit ab.

Atomhülle gebildet aus zwei Membranen – einer äußeren und einer inneren, zwischen denen sich Raum befindet. Der Intermembranraum kommuniziert mit der Höhle des rauen endoplasmatischen Retikulums, und die äußere Membran des Zellkerns kann Ribosomen tragen. Die Kernhülle ist von zahlreichen Poren durchzogen, die mit speziellen Proteinen ausgekleidet sind. Der Transport von Stoffen erfolgt durch die Poren: Die notwendigen Proteine (einschließlich Enzyme), Ionen, Nukleotide und andere Stoffe gelangen in den Kern und RNA-Moleküle, verbrauchte Proteine und Untereinheiten von Ribosomen verlassen ihn. Die Funktionen der Kernhülle bestehen somit in der Trennung des Kerninhalts vom Zytoplasma sowie in der Regulierung des Stoffwechsels zwischen Kern und Zytoplasma.
Nukleoplasma bezeichnet den Inhalt des Zellkerns, in den Chromatin und der Nukleolus eingetaucht sind. Es handelt sich um eine kolloidale Lösung, die chemisch an Zytoplasma erinnert. Enzyme des Nukleoplasmas katalysieren den Austausch von Aminosäuren, Nukleotiden, Proteinen usw. Das Nukleoplasma ist über Kernporen mit dem Hyaloplasma verbunden. Die Funktionen des Nukleoplasmas bestehen ebenso wie des Hyaloplasmas darin, die Verbindung aller Strukturkomponenten des Zellkerns sicherzustellen und eine Reihe enzymatischer Reaktionen durchzuführen.
Chromatin bezeichnet eine Ansammlung dünner Filamente und Körnchen, die im Nukleoplasma eingetaucht sind. Der Nachweis ist nur durch Färbung möglich, da die Brechungsindizes von Chromatin und Nukleoplasma annähernd gleich sind. Der filamentöse Bestandteil des Chromatins wird genannt Euchromatin und körnig - Heterochromatin. Euchromatin ist schwach verdichtet, da daraus Erbinformationen abgelesen werden, während stärker spiralisiertes Heterochromatin genetisch inaktiv ist.
Chromatin ist eine strukturelle Modifikation der Chromosomen in einem sich nicht teilenden Kern. Somit sind Chromosomen ständig im Zellkern vorhanden; nur ihr Zustand ändert sich abhängig von der Funktion, die der Zellkern gerade ausführt.
Die Zusammensetzung des Chromatins umfasst hauptsächlich Nukleoproteinproteine (Desoxyribonukleoproteine und Ribonukleoproteine) sowie Enzyme, von denen die wichtigsten mit der Synthese von Nukleinsäuren verbunden sind, und einige andere Substanzen.
Die Funktionen des Chromatins bestehen erstens in der Synthese von für einen bestimmten Organismus spezifischen Nukleinsäuren, die die Synthese spezifischer Proteine steuern, und zweitens in der Übertragung erblicher Eigenschaften von der Mutterzelle auf die Tochterzellen, wozu die Chromatinfäden werden während des Teilungsprozesses in Chromosomen verpackt.
Nukleolus- ein unter dem Mikroskop deutlich sichtbarer kugelförmiger Körper mit einem Durchmesser von 1-3 Mikrometern. Es wird auf Abschnitten des Chromatins gebildet, in denen Informationen über die Struktur von rRNA und ribosomalen Proteinen kodiert sind. Oft gibt es nur einen Nukleolus im Zellkern, aber in den Zellen, in denen intensive lebenswichtige Prozesse ablaufen, können zwei oder mehr Nukleolen vorhanden sein. Die Funktionen der Nukleolen sind die Synthese von rRNA und der Zusammenbau ribosomaler Untereinheiten durch Kombination von rRNA mit Proteinen, die aus dem Zytoplasma stammen.
Mitochondrien- Doppelmembranorganellen in runder, ovaler oder stäbchenförmiger Form, obwohl auch spiralförmige Organellen vorkommen (in Spermien). Der Durchmesser der Mitochondrien beträgt bis zu 1 µm und die Länge bis zu 7 µm. Der Raum innerhalb der Mitochondrien ist mit Matrix gefüllt. Matrix- Dies ist die Hauptsubstanz der Mitochondrien. Darin sind ein zirkuläres DNA-Molekül und Ribosomen eingetaucht. Die äußere Membran der Mitochondrien ist glatt und für viele Substanzen undurchlässig. Die innere Membran hat Vorsprünge - Cristas, wodurch die Oberfläche der Membranen vergrößert wird, damit chemische Reaktionen ablaufen können. Auf der Oberfläche der Membran befinden sich zahlreiche Proteinkomplexe, die die sogenannte Atmungskette bilden, sowie pilzförmige ATP-Synthetase-Enzyme. In den Mitochondrien findet die aerobe Phase der Atmung statt, in der ATP synthetisiert wird.

Plastiden- große Doppelmembranorganellen, die nur für Pflanzenzellen charakteristisch sind. Der Innenraum der Plastiden ist gefüllt Stroma, oder Matrix. Das Stroma enthält ein mehr oder weniger entwickeltes System von Membranvesikeln - Thylakoide, die in Haufen gesammelt werden - Körner sowie sein eigenes zirkuläres DNA-Molekül und seine Ribosomen. Es gibt vier Haupttypen von Plastiden: Chloroplasten, Chromoplasten, Leukoplasten und Proplastiden.
Chloroplasten- Dies sind grüne Plastiden mit einem Durchmesser von 3-10 Mikrometern, die unter dem Mikroskop deutlich sichtbar sind. Sie kommen nur in den grünen Teilen von Pflanzen vor – Blättern, jungen Stängeln, Blüten und Früchten. Chloroplasten haben im Allgemeinen eine ovale oder ellipsoide Form, können aber auch becherförmig, spiralförmig oder sogar gelappt sein. Die Anzahl der Chloroplasten in einer Zelle beträgt durchschnittlich 10 bis 100 Stück. Bei manchen Algen kann es sich jedoch beispielsweise um eine einzelne Alge handeln, die erhebliche Abmessungen und eine komplexe Form hat – dann heißt es Chromatophor. In anderen Fällen kann die Anzahl der Chloroplasten mehrere Hundert erreichen, während ihre Größe gering ist. Die Farbe der Chloroplasten ist auf das Hauptpigment der Photosynthese zurückzuführen – Chlorophyll, obwohl sie auch zusätzliche Pigmente enthalten - Carotinoide. Carotinoide machen sich erst im Herbst bemerkbar, wenn das Chlorophyll in alternden Blättern abgebaut wird. Die Hauptfunktion von Chloroplasten ist die Photosynthese. Lichtreaktionen der Photosynthese finden auf Thylakoidmembranen statt, an denen Chlorophyllmoleküle befestigt sind, und Dunkelreaktionen finden im Stroma statt, wo zahlreiche Enzyme enthalten sind.

Chromoplasten- Dies sind gelbe, orange und rote Plastiden, die Carotinoidpigmente enthalten. Auch die Form von Chromoplasten kann erheblich variieren: Sie können röhrenförmig, kugelförmig, kristallin usw. sein. Chromoplasten verleihen den Blüten und Früchten von Pflanzen Farbe und locken Bestäuber und Vertreiber von Samen und Früchten an.
Leukoplasten- Dabei handelt es sich um weiße oder farblose Plastiden, meist runder oder ovaler Form. Sie kommen häufig in nicht photosynthetischen Teilen von Pflanzen vor, beispielsweise in der Schale von Blättern, Kartoffelknollen usw. Sie speichern Nährstoffe, am häufigsten Stärke, bei einigen Pflanzen können es sich jedoch auch um Proteine oder Öle handeln.
Plastiden werden in Pflanzenzellen aus Proplastiden gebildet, die bereits in den Zellen des Bildungsgewebes vorhanden sind und kleine Doppelmembrankörper sind. In den frühen Entwicklungsstadien sind verschiedene Arten von Plastiden in der Lage, sich ineinander umzuwandeln: Unter Lichteinwirkung verfärben sich die Leukoplasten einer Kartoffelknolle und die Chromoplasten einer Karottenwurzel grün.
Plastiden und Mitochondrien werden als halbautonome Organellen der Zelle bezeichnet, da sie über eigene DNA-Moleküle und Ribosomen verfügen, die Proteinsynthese durchführen und sich unabhängig von der Zellteilung teilen. Diese Merkmale werden durch ihre Herkunft aus einzelligen prokaryotischen Organismen erklärt. Allerdings ist die „Unabhängigkeit“ von Mitochondrien und Plastiden begrenzt, da ihre DNA zu wenige Gene für eine freie Existenz enthält, während der Rest der Informationen in den Chromosomen des Zellkerns kodiert ist, was es ihm ermöglicht, diese Organellen zu kontrollieren.
Endoplasmatisches Retikulum (ER), oder endoplasmatisches Retikulum (ER) ist eine Einzelmembranorganelle, bei der es sich um ein Netzwerk aus Membranhohlräumen und Tubuli handelt, das bis zu 30 % des Inhalts des Zytoplasmas einnimmt. Der Durchmesser der EPS-Tubuli beträgt etwa 25–30 nm. Es gibt zwei Arten von EPS – raues und glattes. Grobes XPS trägt Ribosomen, in denen die Proteinsynthese stattfindet. Reibungsloses XPS Es fehlen Ribosomen. Seine Funktion ist die Synthese von Lipiden und Kohlenhydraten sowie der Transport, die Speicherung und die Neutralisierung toxischer Substanzen. Es wird vor allem in solchen Zellen entwickelt, in denen intensive Stoffwechselprozesse ablaufen, beispielsweise in Leberzellen – Hepatozyten – und Skelettmuskelfasern. Im ER synthetisierte Substanzen werden zum Golgi-Apparat transportiert. Der Aufbau der Zellmembranen findet ebenfalls im ER statt, ihre Bildung wird jedoch im Golgi-Apparat abgeschlossen.
Golgi-Apparat, oder Golgi-Komplex ist ein einmembraniges Organell, das aus einem System flacher Zisternen, Röhrchen und davon getrennter Bläschen besteht. Die Struktureinheit des Golgi-Apparats ist dictyosome- ein Stapel von Tanks, an dessen einem Pol Substanzen aus dem EPS ankommen und vom gegenüberliegenden Pol nach bestimmten Umwandlungen in Vesikel gepackt und in andere Teile der Zelle geschickt werden. Der Durchmesser der Tanks beträgt etwa 2 Mikrometer und der Durchmesser der kleinen Blasen beträgt etwa 20–30 Mikrometer. Die Hauptfunktionen des Golgi-Komplexes sind die Synthese bestimmter Substanzen und die Modifikation (Veränderung) von Proteinen, Lipiden und Kohlenhydraten aus dem ER, die endgültige Bildung von Membranen sowie der Transport von Substanzen durch die Zelle und die Erneuerung ihrer Strukturen und die Bildung von Lysosomen. Der Golgi-Apparat erhielt seinen Namen zu Ehren des italienischen Wissenschaftlers Camillo Golgi, der dieses Organell erstmals entdeckte (1898).

Lysosomen- kleine Einzelmembranorganellen mit einem Durchmesser von bis zu 1 μm, die hydrolytische Enzyme enthalten, die an der intrazellulären Verdauung beteiligt sind. Die Membranen von Lysosomen sind für diese Enzyme schlecht durchlässig, sodass die Lysosomen ihre Funktionen sehr genau und zielgerichtet erfüllen. Somit nehmen sie aktiv am Prozess der Phagozytose teil, bilden Verdauungsvakuolen und verdauen diese im Falle eines Hungers oder einer Schädigung bestimmter Teile der Zelle, ohne andere zu beeinträchtigen. Die Rolle von Lysosomen bei Zelltodprozessen wurde kürzlich entdeckt.
Vakuole ist ein Hohlraum im Zytoplasma pflanzlicher und tierischer Zellen, der von einer Membran begrenzt und mit Flüssigkeit gefüllt ist. Verdauungs- und kontraktile Vakuolen finden sich in Protozoenzellen. Erstere nehmen am Prozess der Phagozytose teil, indem sie Nährstoffe abbauen. Letztere sorgen durch Osmoregulation für die Aufrechterhaltung des Wasser-Salz-Gleichgewichts. Bei mehrzelligen Tieren kommen hauptsächlich Verdauungsvakuolen vor.
In Pflanzenzellen sind immer Vakuolen vorhanden, sie sind von einer speziellen Membran umgeben und mit Zellsaft gefüllt. Die die Vakuole umgebende Membran ähnelt in ihrer chemischen Zusammensetzung, Struktur und Funktion der Plasmamembran. Zellflüssigkeit ist eine wässrige Lösung verschiedener anorganischer und organischer Substanzen, darunter Mineralsalze, organische Säuren, Kohlenhydrate, Proteine, Glykoside, Alkaloide usw. Die Vakuole kann bis zu 90 % des Zellvolumens einnehmen und den Zellkern an die Peripherie drängen. Dieser Teil der Zelle erfüllt Speicher-, Ausscheidungs-, osmotische, schützende, lysosomale und andere Funktionen, da er Nährstoffe und Abfallprodukte ansammelt, die Wasserversorgung sicherstellt und die Form und das Volumen der Zelle erhält und außerdem Enzyme für den Abbau enthält viele Zellbestandteile. Darüber hinaus können die biologisch aktiven Substanzen der Vakuolen viele Tiere davon abhalten, diese Pflanzen zu fressen. Bei einer Reihe von Pflanzen kommt es aufgrund der Schwellung der Vakuolen zu einem Zellwachstum durch Verlängerung.
Vakuolen sind auch in den Zellen einiger Pilze und Bakterien vorhanden, aber bei Pilzen erfüllen sie nur die Funktion der Osmoregulation, während sie bei Cyanobakterien den Auftrieb aufrechterhalten und an der Aufnahme von Stickstoff aus der Luft beteiligt sind.
Ribosomen- kleine Nichtmembranorganellen mit einem Durchmesser von 15–20 Mikrometern, bestehend aus zwei Untereinheiten – groß und klein. Eukaryotische ribosomale Untereinheiten werden im Nukleolus zusammengesetzt und dann in das Zytoplasma transportiert. Ribosomen in Prokaryoten, Mitochondrien und Plastiden sind kleiner als Ribosomen in Eukaryoten. Ribosomale Untereinheiten umfassen rRNA und Proteine.

Die Anzahl der Ribosomen in einer Zelle kann mehrere zehn Millionen erreichen: Im Zytoplasma, in den Mitochondrien und in den Plastiden befinden sie sich in einem freien Zustand und im rauen ER – in einem gebundenen Zustand. Sie sind an der Proteinsynthese beteiligt, insbesondere führen sie den Translationsprozess durch – die Biosynthese einer Polypeptidkette auf einem mRNA-Molekül. Freie Ribosomen synthetisieren die Proteine von Hyaloplasma, Mitochondrien, Plastiden und ihren eigenen ribosomalen Proteinen, während an das raue ER gebundene Ribosomen die Translation von Proteinen zur Entfernung aus Zellen, den Membranaufbau und die Bildung von Lysosomen und Vakuolen durchführen.
Ribosomen können einzeln im Hyaloplasma gefunden oder während der gleichzeitigen Synthese mehrerer Polypeptidketten auf einer mRNA in Gruppen zusammengesetzt werden. Solche Ribosomengruppen werden genannt Polyribosomen, oder Polysome.

Mikrotubuli- Hierbei handelt es sich um zylindrische hohle Nichtmembranorganellen, die das gesamte Zytoplasma der Zelle durchdringen. Ihr Durchmesser beträgt etwa 25 nm, die Wandstärke beträgt 6–8 nm. Sie werden von zahlreichen Eiweißmolekülen gebildet Tubulin, die zunächst 13 perlenähnliche Fäden bilden und sich dann zu einem Mikrotubulus zusammenfügen. Mikrotubuli bilden ein zytoplasmatisches Retikulum, das der Zelle Form und Volumen verleiht, die Plasmamembran mit anderen Teilen der Zelle verbindet, den Stofftransport durch die Zelle gewährleistet, an der Bewegung der Zelle und intrazellulärer Komponenten sowie an beteiligt ist die Aufteilung des genetischen Materials. Sie sind Teil des Zellzentrums und der Bewegungsorganellen – Flagellen und Zilien.
Mikrofilamente, oder Mikrothreads, sind ebenfalls Nichtmembranorganellen, haben jedoch eine filamentöse Form und werden nicht durch Tubulin, sondern gebildet Aktin. Sie sind an den Prozessen des Membrantransports, der interzellulären Erkennung, der Teilung des Zellzytoplasmas und seiner Bewegung beteiligt. In Muskelzellen vermittelt die Interaktion von Aktin-Mikrofilamenten mit Myosinfilamenten die Kontraktion.
Mikrotubuli und Mikrofilamente bilden das innere Skelett der Zelle – Zytoskelett. Es handelt sich um ein komplexes Fasernetzwerk, das die Plasmamembran mechanisch stützt und die Form der Zelle, die Lage der Zellorganellen und deren Bewegung während der Zellteilung bestimmt.

Zellzentrum- ein Nicht-Membran-Organell, das sich in tierischen Zellen in der Nähe des Zellkerns befindet; es fehlt in Pflanzenzellen. Seine Länge beträgt etwa 0,2–0,3 Mikrometer und sein Durchmesser beträgt 0,1–0,15 Mikrometer. Das Zellzentrum wird von zwei gebildet Zentriolen, in zueinander senkrechten Ebenen liegend, und strahlende Kugel aus Mikrotubuli. Jedes Zentriol besteht aus neun Gruppen von Mikrotubuli, die in Dreiergruppen, d. h. Tripletts, zusammengefasst sind. Das Zellzentrum ist an den Prozessen des Zusammenbaus von Mikrotubuli, der Teilung des Erbmaterials der Zelle sowie an der Bildung von Flagellen und Flimmerhärchen beteiligt.

Bewegungsorganellen. Flagellen Und Zilien Es handelt sich um mit Plasmalemma bedeckte Zellauswüchse. Die Basis dieser Organellen bilden neun Mikrotubulipaare entlang der Peripherie und zwei freie Mikrotubuli in der Mitte. Mikrotubuli sind durch verschiedene Proteine untereinander verbunden und sorgen so für ihre koordinierte Abweichung von der Achse – Schwingung. Schwingungen sind energieabhängig, das heißt, für diesen Prozess wird die Energie hochenergetischer ATP-Bindungen aufgewendet. Die Wiederherstellung verlorener Geißeln und Flimmerhärchen ist eine Funktion Basalkörper, oder Kinetosomen an ihrer Basis gelegen.
Die Länge der Flimmerhärchen beträgt etwa 10–15 nm und die Länge der Flagellen 20–50 µm. Aufgrund der streng gerichteten Bewegungen von Flagellen und Flimmerhärchen erfolgt nicht nur die Bewegung einzelliger Tiere, Spermien usw., sondern auch die Reinigung der Atemwege und die Bewegung der Eizelle durch die Eileiter, da alle diese Teile des menschlichen Körpers sind mit Flimmerepithel ausgekleidet.

Einschlüsse
Einschlüsse- Dabei handelt es sich um instabile Bestandteile der Zelle, die im Laufe ihres Lebens entstehen und wieder verschwinden. Hierzu zählen sowohl Reservestoffe, zum Beispiel Stärke- oder Eiweißkörner in Pflanzenzellen, Glykogenkörnchen in Zellen von Tieren und Pilzen, Volutin in Bakterien, Fetttröpfchen in Zellen aller Art, als auch Abfallprodukte, insbesondere Nahrungsreste Durch die Phagozytose werden sie unverdaut und bilden sogenannte Restkörper.

Die Beziehung zwischen der Struktur und den Funktionen der Teile und Organellen einer Zelle ist die Grundlage ihrer Integrität
Jeder Teil der Zelle ist einerseits eine separate Struktur mit einer bestimmten Struktur und Funktion und andererseits Bestandteil eines komplexeren Systems, das als Zelle bezeichnet wird. Die meisten Erbinformationen einer eukaryotischen Zelle sind im Zellkern konzentriert, der Kern selbst ist jedoch nicht in der Lage, ihre Umsetzung sicherzustellen, da hierfür mindestens das Zytoplasma, das als Hauptsubstanz fungiert, und Ribosomen erforderlich sind, an denen diese Synthese stattfindet . Die meisten Ribosomen befinden sich im granulären endoplasmatischen Retikulum, von wo aus Proteine am häufigsten zum Golgi-Komplex und dann nach Modifikation in die Teile der Zelle transportiert werden, für die sie bestimmt sind oder ausgeschieden werden. Membranverpackungen von Proteinen und Kohlenhydraten können in die Membranen von Organellen und die Zytoplasmamembran eingebettet werden und sorgen so für deren ständige Erneuerung. Auch Lysosomen und Vakuolen, die wichtige Funktionen erfüllen, lösen sich vom Golgi-Komplex. Ohne Lysosomen würden Zellen beispielsweise schnell zu einer Art Mülldeponie für Abfallmoleküle und -strukturen werden.
Der Ablauf all dieser Prozesse erfordert Energie, die von Mitochondrien und in Pflanzen von Chloroplasten produziert wird. Und obwohl diese Organellen relativ autonom sind, da sie über eigene DNA-Moleküle verfügen, werden einige ihrer Proteine immer noch vom Kerngenom kodiert und im Zytoplasma synthetisiert.
Somit ist die Zelle eine untrennbare Einheit ihrer Bestandteile, von denen jeder seine eigene einzigartige Funktion erfüllt.
Stoffwechsel und Energieumwandlung sind Eigenschaften lebender Organismen. Energie- und Kunststoffstoffwechsel, ihre Beziehung. Phasen des Energiestoffwechsels. Gärung und Atmung. Photosynthese, ihre Bedeutung, kosmische Rolle. Phasen der Photosynthese. Helle und dunkle Reaktionen der Photosynthese, ihre Beziehung. Chemosynthese. Die Rolle chemosynthetischer Bakterien auf der Erde
Stoffwechsel und Energieumwandlung – Eigenschaften lebender Organismen
Eine Zelle kann mit einer Miniatur-Chemiefabrik verglichen werden, in der Hunderte und Tausende chemischer Reaktionen ablaufen.
Stoffwechsel- eine Reihe chemischer Transformationen, die auf die Erhaltung und Selbstreproduktion biologischer Systeme abzielen.
Es umfasst die Aufnahme von Stoffen in den Körper während der Ernährung und Atmung, des intrazellulären Stoffwechsels oder Stoffwechsel sowie die Isolierung der Stoffwechselendprodukte.
Der Stoffwechsel ist untrennbar mit den Prozessen der Umwandlung einer Energieart in eine andere verbunden. Während des Prozesses der Photosynthese wird beispielsweise Lichtenergie in Form der Energie chemischer Bindungen komplexer organischer Moleküle gespeichert und während des Atmungsprozesses freigesetzt und für die Synthese neuer Moleküle sowie für mechanische und osmotische Arbeit aufgewendet. in Form von Wärme etc. abgegeben werden.
Das Auftreten chemischer Reaktionen in lebenden Organismen wird durch biologische Katalysatoren mit Proteincharakter sichergestellt - Enzyme, oder Enzyme. Wie andere Katalysatoren beschleunigen Enzyme den Ablauf chemischer Reaktionen in einer Zelle um das Zehn- und Hunderttausendfache und machen sie manchmal sogar möglich, verändern aber weder die Natur noch die Eigenschaften des Endprodukts bzw. der Endprodukte der Reaktion und tun dies auch sich nicht ändern. Enzyme können sowohl einfache als auch komplexe Proteine sein, die neben dem Proteinanteil auch einen Nicht-Proteinanteil enthalten – Cofaktor (Coenzym). Beispiele für Enzyme sind Speichelamylase, die bei längerem Kauen Polysaccharide abbaut, und Pepsin, das für die Verdauung von Proteinen im Magen sorgt.
Enzyme unterscheiden sich von Nicht-Protein-Katalysatoren durch ihre hohe Wirkungsspezifität, eine deutliche Steigerung der Reaktionsgeschwindigkeit mit ihrer Hilfe sowie die Fähigkeit, die Wirkung durch Veränderung der Reaktionsbedingungen oder die Wechselwirkung verschiedener Stoffe mit ihnen zu regulieren. Darüber hinaus unterscheiden sich die Bedingungen, unter denen die enzymatische Katalyse stattfindet, erheblich von denen, unter denen die nicht-enzymatische Katalyse stattfindet: Die optimale Temperatur für die Funktion von Enzymen im menschlichen Körper beträgt 37 °C, der Druck sollte nahe am Atmosphärendruck liegen und die Der pH-Wert der Umgebung kann erheblich schwanken. Daher benötigt Amylase ein alkalisches Milieu und Pepsin ein saures Milieu.
Der Wirkungsmechanismus von Enzymen besteht darin, die Aktivierungsenergie von Substanzen (Substraten) zu reduzieren, die durch die Bildung intermediärer Enzym-Substrat-Komplexe eine Reaktion eingehen.

Energie- und Kunststoffstoffwechsel, ihre Beziehung
Der Stoffwechsel besteht aus zwei Prozessen, die gleichzeitig in der Zelle ablaufen: dem Kunststoff- und dem Energiestoffwechsel.
Plastischer Stoffwechsel (Anabolismus, Assimilation) ist eine Reihe von Synthesereaktionen, bei denen ATP-Energie verbraucht wird. Im Prozess des plastischen Stoffwechsels werden für die Zelle notwendige organische Substanzen synthetisiert. Beispiele für plastische Austauschreaktionen sind Photosynthese, Proteinbiosynthese und DNA-Replikation (Selbstverdopplung).
Energiestoffwechsel (Katabolismus, Dissimilation) ist eine Reihe von Reaktionen, die komplexe Substanzen in einfachere zerlegen. Durch den Energiestoffwechsel wird Energie freigesetzt und in Form von ATP gespeichert. Die wichtigsten Prozesse des Energiestoffwechsels sind Atmung und Fermentation.
Kunststoff- und Energieaustausch sind untrennbar miteinander verbunden, da im Prozess des Kunststoffaustauschs organische Substanzen synthetisiert werden und dafür ATP-Energie benötigt wird und im Prozess des Energieaustauschs organische Substanzen abgebaut und Energie freigesetzt wird, die dann für Syntheseprozesse aufgewendet wird .
Organismen nehmen Energie während des Ernährungsprozesses auf und geben sie hauptsächlich während des Atmungsprozesses ab und wandeln sie in eine zugängliche Form um. Je nach Ernährungsweise werden alle Organismen in Autotrophe und Heterotrophe unterteilt. Autotrophe in der Lage, selbstständig organische Substanzen aus anorganischen zu synthetisieren, und Heterotrophe Verwenden Sie ausschließlich aufbereitete organische Substanzen.
Phasen des Energiestoffwechsels
Trotz der Komplexität der Energiestoffwechselreaktionen werden sie herkömmlicherweise in drei Phasen unterteilt: vorbereitend, anaerob (sauerstofffrei) und aerob (sauerstofffrei).
An Vorbereitungsphase Moleküle aus Polysacchariden, Lipiden, Proteinen und Nukleinsäuren zerfallen in einfachere Moleküle, zum Beispiel Glukose, Glycerin und Fettsäuren, Aminosäuren, Nukleotide usw. Dieses Stadium kann direkt in den Zellen oder im Darm stattfinden, von wo aus sie abgebaut werden Daunenstoffe werden über den Blutkreislauf abgegeben.
Anaerobes Stadium Der Energiestoffwechsel geht mit einem weiteren Abbau der Monomere organischer Verbindungen in noch einfachere Zwischenprodukte einher, beispielsweise Brenztraubensäure oder Pyruvat. Es erfordert keinen Sauerstoff und ist für viele Organismen, die im Schlamm von Sümpfen oder im menschlichen Darm leben, die einzige Möglichkeit, Energie zu gewinnen. Die anaerobe Phase des Energiestoffwechsels findet im Zytoplasma statt.
Verschiedene Stoffe können sauerstofffrei gespalten werden, das Substrat der Reaktionen ist jedoch häufig Glucose. Der Prozess seiner sauerstofffreien Spaltung wird aufgerufen Glykolyse. Bei der Glykolyse verliert ein Glucosemolekül vier Wasserstoffatome, d. h. es wird oxidiert, und es entstehen zwei Moleküle Brenztraubensäure, zwei Moleküle ATP und zwei Moleküle des reduzierten Wasserstoffträgers $NADH + H^(+)$:
$C_6H_(12)O_6 + 2H_3PO_4 + 2ADP + 2NAD → 2C_3H_4O_3 + 2ATP + 2NADH + H^(+) + 2H_2O$.
Die Bildung von ATP aus ADP erfolgt durch die direkte Übertragung von Phosphatanionen aus vorphosphoryliertem Zucker und wird aufgerufen Substratphosphorylierung.
Aerobic-Stufe Der Energieaustausch kann nur in Gegenwart von Sauerstoff stattfinden, während bei der sauerstofffreien Spaltung gebildete Zwischenverbindungen zu den Endprodukten (Kohlendioxid und Wasser) oxidiert werden und der größte Teil der in den chemischen Bindungen organischer Verbindungen gespeicherten Energie freigesetzt wird. Es wird in die Energie hochenergetischer Bindungen von 36 ATP-Molekülen umgewandelt. Diese Stufe wird auch genannt Gewebeatmung. In Abwesenheit von Sauerstoff werden Zwischenverbindungen in andere organische Substanzen umgewandelt, ein sogenannter Prozess Fermentation.
Atem
Der Mechanismus der Zellatmung ist in Abb. schematisch dargestellt.

Aerobe Atmung findet in Mitochondrien statt, wobei Brenztraubensäure zunächst ein Kohlenstoffatom verliert, was mit der Synthese eines reduzierenden Äquivalents von $NADH + H^(+)$ und eines Moleküls Acetyl-Coenzym A (Acetyl-CoA) einhergeht:
$C_3H_4O_3 + NAD + H~CoA → CH_3CO~CoA + NADH + H^(+) + CO_2$.
Acetyl-CoA in der mitochondrialen Matrix ist an einer Kette chemischer Reaktionen beteiligt, deren Gesamtheit aufgerufen wird Krebs Zyklus (Tricarbonsäurezyklus, Zitronensäurezyklus). Bei diesen Umwandlungen entstehen zwei ATP-Moleküle, Acetyl-CoA wird vollständig zu Kohlendioxid oxidiert und seine Wasserstoffionen und Elektronen werden an die Wasserstoffträger $NADH + H^(+)$ und $FADH_2$ addiert. Die Träger transportieren Wasserstoffprotonen und Elektronen zu den inneren Membranen der Mitochondrien und bilden dort Cristae. Mit Hilfe von Trägerproteinen werden Wasserstoffprotonen in den Zwischenmembranraum gepumpt und Elektronen über die sogenannte Atmungskette von Enzymen, die sich auf der Innenmembran der Mitochondrien befinden, übertragen und an Sauerstoffatome abgegeben:
$O_2+2e^(-)→O_2^-$.
Es ist zu beachten, dass einige Proteine der Atmungskette Eisen und Schwefel enthalten.
Aus dem Intermembranraum werden Wasserstoffprotonen mit Hilfe spezieller Enzyme – ATP-Synthasen – zurück in die mitochondriale Matrix transportiert und die dabei freigesetzte Energie für die Synthese von 34 ATP-Molekülen aus jedem Glucosemolekül aufgewendet. Dieser Vorgang wird aufgerufen oxidative Phosphorylierung. In der mitochondrialen Matrix reagieren Wasserstoffprotonen mit Sauerstoffradikalen unter Bildung von Wasser:
$4H^(+)+O_2^-→2H_2O$.
Der Reaktionssatz der Sauerstoffatmung kann wie folgt ausgedrückt werden:
$2C_3H_4O_3 + 6O_2 + 36H_3PO_4 + 36ADP → 6CO_2 + 38H_2O + 36ATP.$
Die allgemeine Atemgleichung sieht folgendermaßen aus:
$C_6H_(12)O_6 + 6O_2 + 38H_3PO_4 + 38ADP → 6CO_2 + 40H_2O + 38ATP.$
Fermentation
In Abwesenheit oder Mangel an Sauerstoff kommt es zur Gärung. Die Fermentation ist eine evolutionär frühere Methode der Energiegewinnung als die Atmung, sie ist jedoch energetisch weniger vorteilhaft, da durch die Fermentation noch energiereiche organische Stoffe entstehen. Es gibt mehrere Haupttypen der Fermentation: Milchsäure, Alkoholsäure, Essigsäure usw. So wird in der Skelettmuskulatur in Abwesenheit von Sauerstoff während der Fermentation Brenztraubensäure zu Milchsäure reduziert, während die zuvor gebildeten Reduktionsäquivalente verbraucht werden, und zwar nur Es bleiben zwei ATP-Moleküle übrig:
$2C_3H_4O_3 + 2NADH + H^(+) → 2C_3H_6O_3 + 2NAD$.
Bei der Gärung mit Hilfe von Hefe wird Brenztraubensäure in Gegenwart von Sauerstoff in Ethylalkohol und Kohlenmonoxid (IV) umgewandelt:
$C_3H_4O_3 + NADH + H^(+) → C_2H_5OH + CO_2 + NAD^(+)$.
Bei der Gärung mit Hilfe von Mikroorganismen können aus Brenztraubensäure auch Essigsäure, Buttersäure, Ameisensäure etc. entstehen.
ATP, das durch den Energiestoffwechsel gewonnen wird, wird in der Zelle für verschiedene Arten von Arbeiten verbraucht: chemische, osmotische, elektrische, mechanische und regulatorische. Chemische Arbeit umfasst die Biosynthese von Proteinen, Lipiden, Kohlenhydraten, Nukleinsäuren und anderen lebenswichtigen Verbindungen. Osmotische Arbeit umfasst die Prozesse der Aufnahme und Entfernung von Substanzen durch die Zelle, die sich im extrazellulären Raum in höheren Konzentrationen als in der Zelle selbst befinden. Elektrische Arbeit steht in engem Zusammenhang mit osmotischer Arbeit, da durch die Bewegung geladener Teilchen durch Membranen eine Membranladung entsteht und die Eigenschaften Erregbarkeit und Leitfähigkeit erworben werden. Bei der mechanischen Arbeit handelt es sich um die Bewegung von Substanzen und Strukturen innerhalb der Zelle sowie der Zelle als Ganzes. Zur regulatorischen Arbeit gehören alle Prozesse, die darauf abzielen, Prozesse in der Zelle zu koordinieren.
Photosynthese, ihre Bedeutung, kosmische Rolle
Photosynthese ist der Prozess der Umwandlung von Lichtenergie in die Energie chemischer Bindungen organischer Verbindungen unter Beteiligung von Chlorophyll.
Durch die Photosynthese werden jährlich etwa 150 Milliarden Tonnen organisches Material und etwa 200 Milliarden Tonnen Sauerstoff produziert. Dieser Prozess stellt den Kohlenstoffkreislauf in der Biosphäre sicher, verhindert die Ansammlung von Kohlendioxid und verhindert so den Treibhauseffekt und die Überhitzung der Erde. Bei der Photosynthese entstehende organische Stoffe werden von anderen Organismen nicht vollständig verbraucht, ein erheblicher Teil davon hat im Laufe der Jahrmillionen Ablagerungen von Mineralien (Stein- und Braunkohle, Erdöl) gebildet. Seit Kurzem werden auch Rapsöl („Biodiesel“) und aus Pflanzenresten gewonnener Alkohol als Treibstoff eingesetzt. Unter dem Einfluss elektrischer Entladungen entsteht aus Sauerstoff Ozon, das einen Ozonschirm bildet, der alles Leben auf der Erde vor den zerstörerischen Auswirkungen ultravioletter Strahlen schützt.
Unser Landsmann, der herausragende Pflanzenphysiologe K. A. Timiryazev (1843-1920), nannte die Rolle der Photosynthese „kosmisch“, da sie die Erde mit der Sonne (dem Weltraum) verbindet und so für einen Energiezufluss zum Planeten sorgt.
Phasen der Photosynthese. Helle und dunkle Reaktionen der Photosynthese, ihre Beziehung
Im Jahr 1905 entdeckte der englische Pflanzenphysiologe F. Blackman, dass die Photosyntheserate nicht unbegrenzt steigen kann, sondern durch einen Faktor begrenzt wird. Darauf aufbauend stellte er die Hypothese auf, dass es zwei Phasen der Photosynthese gibt: Licht Und dunkel. Bei geringer Lichtintensität erhöht sich die Geschwindigkeit der Lichtreaktionen proportional zur Zunahme der Lichtintensität. Darüber hinaus sind diese Reaktionen nicht von der Temperatur abhängig, da für das Auftreten keine Enzyme erforderlich sind. An Thylakoidmembranen treten Lichtreaktionen auf.
Im Gegensatz dazu nimmt die Geschwindigkeit der Dunkelreaktionen mit steigender Temperatur zu. Bei Erreichen einer Temperaturschwelle von 30 °C hört dieser Anstieg jedoch auf, was auf die enzymatische Natur dieser im Stroma stattfindenden Umwandlungen hinweist. Es ist zu beachten, dass Licht auch einen gewissen Einfluss auf Dunkelreaktionen hat, obwohl diese als Dunkelreaktionen bezeichnet werden.

Die Lichtphase der Photosynthese findet auf Thylakoidmembranen statt, die verschiedene Arten von Proteinkomplexen tragen, von denen die Photosysteme I und II sowie die ATP-Synthase die wichtigsten sind. Zu den Photosystemen zählen Pigmentkomplexe, die neben Chlorophyll auch Carotinoide enthalten. Carotinoide fangen Licht in Bereichen des Spektrums ein, in denen Chlorophyll dies nicht tut, und schützen Chlorophyll außerdem vor der Zerstörung durch hochintensives Licht.
Zu den Photosystemen gehören neben Pigmentkomplexen auch eine Reihe von Elektronenakzeptorproteinen, die nacheinander Elektronen von Chlorophyllmolekülen aufeinander übertragen. Die Reihenfolge dieser Proteine wird aufgerufen Elektronentransportkette von Chloroplasten.
Mit dem Photosystem II ist auch ein spezieller Proteinkomplex verbunden, der für die Freisetzung von Sauerstoff bei der Photosynthese sorgt. Dieser Sauerstoff freisetzende Komplex enthält Mangan- und Chlorionen.
IN Lichtphase Lichtquanten oder Photonen, die auf Chlorophyllmoleküle fallen, die sich auf Thylakoidmembranen befinden, überführen sie in einen angeregten Zustand, der durch eine höhere Elektronenenergie gekennzeichnet ist. In diesem Fall werden angeregte Elektronen aus dem Chlorophyll des Photosystems I über eine Kette von Vermittlern auf den Wasserstoffträger NADP übertragen, der Wasserstoffprotonen anlagert, die in einer wässrigen Lösung immer vorhanden sind:
$NADP + 2e^(-) + 2H^(+) → NADPH + H^(+)$.
Das reduzierte $NADPH + H^(+)$ wird anschließend im Dunkelstadium verwendet. Elektronen aus dem Chlorophyll des Photosystems II werden ebenfalls entlang der Elektronentransportkette übertragen, sie füllen jedoch die „Elektronenlöcher“ des Chlorophylls des Photosystems I. Der Mangel an Elektronen im Chlorophyll des Photosystems II wird durch den Entzug von Wassermolekülen ausgeglichen, die erfolgt unter Beteiligung des oben bereits erwähnten Sauerstoff freisetzenden Komplexes. Infolge der Zersetzung von Wassermolekülen, die man nennt Photolyse Es entstehen Wasserstoffprotonen und es wird molekularer Sauerstoff freigesetzt, der ein Nebenprodukt der Photosynthese ist:
$H_2O → 2H^(+) + 2e^(-) + (1)/(2)O_2$.
Genetische Informationen in einer Zelle. Gene, genetischer Code und seine Eigenschaften. Matrixnatur von Biosynthesereaktionen. Biosynthese von Proteinen und Nukleinsäuren
Genetische Informationen in einer Zelle
Die Fortpflanzung unter Artgenossen ist eine der grundlegenden Eigenschaften von Lebewesen. Dank dieses Phänomens gibt es nicht nur Ähnlichkeiten zwischen Organismen, sondern auch zwischen einzelnen Zellen sowie ihren Organellen (Mitochondrien und Plastiden). Die materielle Grundlage dieser Ähnlichkeit ist die in der DNA-Nukleotidsequenz verschlüsselte Übertragung genetischer Informationen, die durch Prozesse der DNA-Replikation (Selbstverdopplung) erfolgt. Alle Eigenschaften und Eigenschaften von Zellen und Organismen werden durch Proteine verwirklicht, deren Struktur hauptsächlich durch die Sequenz der DNA-Nukleotide bestimmt wird. Daher kommt der Biosynthese von Nukleinsäuren und Proteinen bei Stoffwechselprozessen eine herausragende Bedeutung zu. Die Struktureinheit der Erbinformation ist das Gen.
Gene, genetischer Code und seine Eigenschaften
Erbinformationen in einer Zelle sind nicht monolithisch, sondern in einzelne „Wörter“ – Gene – unterteilt.
Gen ist eine elementare Einheit der genetischen Information.
Die Arbeit am „Human Genome“-Programm, das gleichzeitig in mehreren Ländern durchgeführt und zu Beginn dieses Jahrhunderts abgeschlossen wurde, brachte uns die Erkenntnis, dass ein Mensch nur über etwa 25.000 bis 30.000 Gene verfügt, aber Informationen aus dem Großteil unserer DNA wird nie gelesen, da es eine große Anzahl bedeutungsloser Abschnitte, Wiederholungen und Gene enthält, die Merkmale kodieren, die für den Menschen ihre Bedeutung verloren haben (Schwanz, Körperbehaarung usw.). Darüber hinaus wurden eine Reihe von Genen entschlüsselt, die für die Entstehung von Erbkrankheiten verantwortlich sind, sowie Zielgene für Medikamente. Die praktische Anwendung der bei der Umsetzung dieses Programms erzielten Ergebnisse wird jedoch verschoben, bis die Genome weiterer Menschen entschlüsselt sind und klar wird, wie sie sich unterscheiden.
Gene, die die Primärstruktur von Proteinen, Ribosomen oder Transfer-RNA kodieren, werden als Gene bezeichnet strukturell und Gene, die für die Aktivierung oder Unterdrückung von Leseinformationen aus Strukturgenen sorgen – regulatorisch. Allerdings enthalten auch Strukturgene regulatorische Regionen.
Die Erbinformationen von Organismen sind in Form bestimmter Nukleotidkombinationen und ihrer Sequenz in der DNA verschlüsselt – genetischer Code. Seine Eigenschaften sind: Triplettität, Spezifität, Universalität, Redundanz und Nichtüberlappung. Darüber hinaus gibt es im genetischen Code keine Satzzeichen.
Jede Aminosäure wird in der DNA durch drei Nukleotide kodiert – Triplett, Beispielsweise wird Methionin durch das TAC-Triplett kodiert, das heißt, der Code ist ein Triplett. Andererseits kodiert jedes Triplett nur eine Aminosäure, was seine Spezifität oder Eindeutigkeit ausmacht. Der genetische Code ist für alle lebenden Organismen universell, das heißt, Erbinformationen über menschliche Proteine können von Bakterien gelesen werden und umgekehrt. Dies weist auf die Einheit des Ursprungs der organischen Welt hin. Allerdings entsprechen 64 Kombinationen von drei Nukleotiden nur 20 Aminosäuren, wodurch eine Aminosäure durch 2-6 Tripletts kodiert werden kann, das heißt, der genetische Code ist redundant oder degeneriert. Drei Tripletts haben keine entsprechenden Aminosäuren, sie werden genannt Stopp-Codons, da sie das Ende der Synthese der Polypeptidkette anzeigen.
Die Basensequenz in DNA-Tripletts und die von ihnen kodierten Aminosäuren

*Stoppcodon, das das Ende der Synthese der Polypeptidkette anzeigt.
Abkürzungen für Aminosäurenamen:
Ala – Alanin
Arg – Arginin
Asn – Asparagin
Asp – Asparaginsäure
Val - Valin
Sein - Histidin
Gly – Glycin
Gln – Glutamin
Glu – Glutaminsäure
Ile – Isoleucin
Leu - Leucin
Liz - Lysin
Meth – Methionin
Pro - Prolin
Ser - Serin
Tyr – Tyrosin
Tre – Threonin
Drei – Tryptophan
Fen - Phenylalanin
Cis – Cystein
Wenn Sie beginnen, genetische Informationen nicht vom ersten Nukleotid im Triplett abzulesen, sondern vom zweiten, dann verschiebt sich nicht nur der Leserahmen, sondern das auf diese Weise synthetisierte Protein wird nicht nur in der Nukleotidsequenz, sondern auch in völlig anders sein Struktur und Eigenschaften. Es gibt keine Satzzeichen zwischen den Tripletts, sodass der Verschiebung des Leserahmens keine Hindernisse entgegenstehen, was Raum für das Auftreten und die Aufrechterhaltung von Mutationen eröffnet.
Matrixnatur von Biosynthesereaktionen
Bakterienzellen können sich alle 20 bis 30 Minuten verdoppeln, eukaryotische Zellen jeden Tag und sogar noch häufiger, was eine hohe Geschwindigkeit und Genauigkeit der DNA-Replikation erfordert. Darüber hinaus enthält jede Zelle Hunderte und Tausende Kopien vieler Proteine, insbesondere Enzyme, weshalb die „stückweise“ Methode ihrer Produktion für ihre Reproduktion inakzeptabel ist. Eine fortschrittlichere Methode ist das Stempeln, mit dem Sie zahlreiche exakte Kopien des Produkts erhalten und gleichzeitig dessen Kosten senken können. Zum Stempeln wird eine Matrize benötigt, aus der der Abdruck hergestellt wird.
In Zellen besteht das Prinzip der Template-Synthese darin, dass neue Moleküle von Proteinen und Nukleinsäuren gemäß dem Programm synthetisiert werden, das in die Struktur bereits vorhandener Moleküle derselben Nukleinsäuren (DNA oder RNA) eingebettet ist.
Biosynthese von Proteinen und Nukleinsäuren
DNA Replikation. DNA ist ein doppelsträngiges Biopolymer, dessen Monomere Nukleotide sind. Wenn die DNA-Biosynthese nach dem Prinzip der Fotokopie erfolgen würde, würde es unweigerlich zu zahlreichen Verzerrungen und Fehlern in der Erbinformation kommen, die letztendlich zum Tod neuer Organismen führen würden. Daher läuft der Prozess der DNA-Verdoppelung anders ab, auf halbkonservative Weise: Das DNA-Molekül entwindet sich und an jeder der Ketten wird nach dem Prinzip der Komplementarität eine neue Kette synthetisiert. Der Prozess der Selbstreproduktion eines DNA-Moleküls, der eine genaue Kopie der Erbinformationen und deren Weitergabe von Generation zu Generation gewährleistet, wird als bezeichnet Reproduzieren(von lat. Replikation- Wiederholung). Durch die Replikation entstehen zwei absolut exakte Kopien des Mutter-DNA-Moleküls, von denen jede eine Kopie des Mutter-DNA-Moleküls trägt.

Der Replikationsprozess ist tatsächlich äußerst komplex, da eine Reihe von Proteinen daran beteiligt sind. Einige von ihnen wickeln die Doppelhelix der DNA ab, andere brechen die Wasserstoffbrückenbindungen zwischen den Nukleotiden komplementärer Ketten, andere (zum Beispiel das Enzym DNA-Polymerase) wählen neue Nukleotide nach dem Prinzip der Komplementarität aus usw. Zwei DNA-Moleküle bilden sich als a Das Ergebnis der Replikation zerfällt in zwei bei der Teilung neu gebildete Tochterzellen.
Fehler im Replikationsprozess treten äußerst selten auf, wenn sie aber dennoch auftreten, werden sie sowohl durch DNA-Polymerasen als auch durch spezielle Reparaturenzyme sehr schnell beseitigt, da jeder Fehler in der Nukleotidsequenz zu einer irreversiblen Veränderung der Struktur und Funktion des Proteins führen kann und letztendlich die Lebensfähigkeit einer neuen Zelle oder sogar eines Individuums beeinträchtigen.
Proteinbiosynthese. Wie der herausragende Philosoph des 19. Jahrhunderts F. Engels es im übertragenen Sinne ausdrückte: „Leben ist eine Existenzform von Proteinkörpern.“ Die Struktur und Eigenschaften von Proteinmolekülen werden durch ihre Primärstruktur bestimmt, d. h. die Sequenz der in der DNA kodierten Aminosäuren. Nicht nur die Existenz des Polypeptids selbst, sondern auch das Funktionieren der Zelle als Ganzes hängt von der Genauigkeit der Reproduktion dieser Informationen ab, daher ist der Prozess der Proteinsynthese von großer Bedeutung. Es scheint der komplexeste Syntheseprozess in der Zelle zu sein, da bis zu dreihundert verschiedene Enzyme und andere Makromoleküle beteiligt sind. Zudem fließt es mit hoher Geschwindigkeit, was eine noch höhere Präzision erfordert.
Es gibt zwei Hauptstadien der Proteinbiosynthese: Transkription und Translation.
Transkription(von lat. Transkription- Umschreiben) ist die Biosynthese von mRNA-Molekülen auf einer DNA-Matrix.

Da das DNA-Molekül zwei antiparallele Ketten enthält, würde das Lesen von Informationen aus beiden Ketten zur Bildung völlig unterschiedlicher mRNAs führen, daher ist ihre Biosynthese nur an einer der Ketten möglich, die im Gegensatz zur zweiten als kodierend oder kodogen bezeichnet wird. nicht-kodierend oder nicht-kodogen. Für den Umschreibvorgang sorgt ein spezielles Enzym, die RNA-Polymerase, das RNA-Nukleotide nach dem Prinzip der Komplementarität auswählt. Dieser Prozess kann sowohl im Zellkern als auch in Organellen mit eigener DNA – Mitochondrien und Plastiden – ablaufen.
Die während der Transkription synthetisierten mRNA-Moleküle durchlaufen einen komplexen Vorbereitungsprozess für die Translation (mitochondriale und plastidäre mRNAs können in den Organellen verbleiben, wo die zweite Stufe der Proteinbiosynthese stattfindet). Während des Prozesses der mRNA-Reifung werden die ersten drei Nukleotide (AUG) und ein Schwanz aus Adenylnukleotiden daran befestigt, deren Länge bestimmt, wie viele Kopien des Proteins auf einem bestimmten Molekül synthetisiert werden können. Erst dann verlassen reife mRNAs den Zellkern durch Kernporen.
Parallel dazu findet im Zytoplasma der Prozess der Aminosäureaktivierung statt, bei dem sich die Aminosäure mit der entsprechenden freien tRNA verbindet. Dieser Prozess wird durch ein spezielles Enzym katalysiert und erfordert ATP.
Übertragen(von lat. übertragen- Transfer) ist die Biosynthese einer Polypeptidkette auf einer mRNA-Matrix, bei der genetische Informationen in die Aminosäuresequenz der Polypeptidkette übersetzt werden.

Die zweite Stufe der Proteinsynthese findet am häufigsten im Zytoplasma statt, beispielsweise im rauen ER. Für sein Auftreten sind die Anwesenheit von Ribosomen, die Aktivierung der tRNA, bei der sie die entsprechenden Aminosäuren anlagern, die Anwesenheit von Mg2+-Ionen sowie optimale Umgebungsbedingungen (Temperatur, pH-Wert, Druck usw.) notwendig.
Um mit der Übertragung zu beginnen ( Einleitung) Eine kleine ribosomale Untereinheit wird an ein zur Synthese bereites mRNA-Molekül gebunden und anschließend wird nach dem Prinzip der Komplementarität zum ersten Codon (AUG) eine tRNA ausgewählt, die die Aminosäure Methionin trägt. Erst danach heftet sich die große ribosomale Untereinheit an. Innerhalb des zusammengesetzten Ribosoms gibt es zwei mRNA-Codons, von denen das erste bereits besetzt ist. Eine zweite tRNA, die ebenfalls eine Aminosäure trägt, wird an das angrenzende Codon angefügt, woraufhin mithilfe von Enzymen eine Peptidbindung zwischen den Aminosäureresten gebildet wird. Das Ribosom bewegt ein Codon der mRNA; Die erste von einer Aminosäure befreite tRNA kehrt nach der nächsten Aminosäure in das Zytoplasma zurück und an der verbleibenden tRNA hängt sozusagen ein Fragment der zukünftigen Polypeptidkette. Die nächste tRNA wird an das neue Codon, das sich innerhalb des Ribosoms befindet, angehängt, der Vorgang wiederholt sich und Schritt für Schritt verlängert sich die Polypeptidkette, d. h. Verlängerung.
Ende der Proteinsynthese ( Beendigung) tritt auf, sobald im mRNA-Molekül eine bestimmte Nukleotidsequenz angetroffen wird, die nicht für eine Aminosäure kodiert (Stoppcodon). Danach werden Ribosom, mRNA und Polypeptidkette getrennt, und das neu synthetisierte Protein erhält die entsprechende Struktur und wird zu dem Teil der Zelle transportiert, wo es seine Funktionen erfüllen wird.
Die Translation ist ein sehr energieintensiver Prozess, da die Energie eines ATP-Moleküls verbraucht wird, um eine Aminosäure an die tRNA zu binden, und mehrere weitere verwendet werden, um das Ribosom entlang des mRNA-Moleküls zu bewegen.
Um die Synthese bestimmter Proteinmoleküle zu beschleunigen, können nacheinander mehrere Ribosomen an ein mRNA-Molekül gebunden werden, die eine einzige Struktur bilden – Polysom.
Eine Zelle ist die genetische Einheit eines Lebewesens. Chromosomen, ihre Struktur (Form und Größe) und Funktionen. Die Anzahl der Chromosomen und ihre Artenkonstanz. Somatische und Keimzellen. Zelllebenszyklus: Interphase und Mitose. Mitose ist die Teilung somatischer Zellen. Meiose. Phasen der Mitose und Meiose. Entwicklung von Keimzellen bei Pflanzen und Tieren. Die Zellteilung ist die Grundlage für das Wachstum, die Entwicklung und die Fortpflanzung von Organismen. Die Rolle von Meiose und Mitose
Eine Zelle ist die genetische Einheit eines Lebewesens.
Obwohl Nukleinsäuren Träger genetischer Informationen sind, ist die Umsetzung dieser Informationen außerhalb der Zelle unmöglich, was am Beispiel von Viren leicht nachgewiesen werden kann. Diese Organismen, die oft nur DNA oder RNA enthalten, können sich nicht selbstständig vermehren; dazu müssen sie den Erbapparat der Zelle nutzen. Ohne die Hilfe der Zelle selbst können sie nicht einmal in eine Zelle eindringen, außer durch den Einsatz von Membrantransportmechanismen oder aufgrund einer Zellschädigung. Die meisten Viren sind instabil; sie sterben bereits nach wenigen Stunden im Freien ab. Folglich ist eine Zelle eine genetische Einheit eines Lebewesens, die über einen Mindestsatz an Komponenten zur Erhaltung, Veränderung und Umsetzung erblicher Informationen sowie deren Weitergabe an Nachkommen verfügt.
Die meisten genetischen Informationen einer eukaryotischen Zelle befinden sich im Zellkern. Die Besonderheit seiner Organisation besteht darin, dass die DNA-Moleküle von Eukaryoten im Gegensatz zur DNA einer prokaryotischen Zelle nicht geschlossen sind und mit Proteinen – Chromosomen – komplexe Komplexe bilden.
Chromosomen, ihre Struktur (Form und Größe) und Funktionen
Chromosom(aus dem Griechischen Chrom- Farbe, Färbung und soma- Körper) ist die Struktur des Zellkerns, der Gene enthält und bestimmte erbliche Informationen über die Eigenschaften und Eigenschaften des Organismus trägt.
Manchmal werden die zirkulären DNA-Moleküle von Prokaryoten auch Chromosomen genannt. Chromosomen sind zur Selbstvervielfältigung fähig; sie besitzen strukturelle und funktionelle Individualität und behalten diese über Generationen hinweg. Jede Zelle trägt alle Erbinformationen des Körpers, aber nur ein kleiner Teil arbeitet in ihr.
Die Basis eines Chromosoms ist ein doppelsträngiges DNA-Molekül, das mit Proteinen gefüllt ist. Bei Eukaryoten interagieren Histon- und Nicht-Histon-Proteine mit der DNA, wohingegen bei Prokaryoten Histon-Proteine fehlen.
Unter dem Lichtmikroskop erkennt man Chromosomen am besten während der Zellteilung, wenn sie durch die Verdichtung das Aussehen von stäbchenförmigen Körpern annehmen, die durch eine primäre Verengung getrennt sind – Zentromer — auf den Schultern. Auf einem Chromosom kann es auch sein sekundäre Verengung, was in manchen Fällen das sogenannte trennt Satellit. Die Enden der Chromosomen werden genannt Telomere. Telomere verhindern das Zusammenkleben der Chromosomenenden und sorgen für deren Befestigung an der Kernmembran in einer sich nicht teilenden Zelle. Zu Beginn der Teilung verdoppeln sich die Chromosomen und bestehen aus zwei Tochterchromosomen – Chromatid, am Zentromer befestigt.

Entsprechend ihrer Form werden Chromosomen in gleicharmige, ungleicharmige und stäbchenförmige Chromosomen unterteilt. Die Größe der Chromosomen variiert erheblich, aber das durchschnittliche Chromosom hat Abmessungen von 5 x 1,4 Mikrometern.
In einigen Fällen enthalten Chromosomen aufgrund zahlreicher DNA-Duplikationen Hunderte und Tausende von Chromatiden: Solche Riesenchromosomen werden genannt Polyäthylen. Man findet sie in den Speicheldrüsen von Drosophila-Larven sowie in den Verdauungsdrüsen von Spulwürmern.
Die Anzahl der Chromosomen und ihre Artenkonstanz. Somatische und Keimzellen
Nach der Zelltheorie ist eine Zelle eine Einheit der Struktur, Lebenstätigkeit und Entwicklung eines Organismus. Somit werden so wichtige Funktionen von Lebewesen wie Wachstum, Fortpflanzung und Entwicklung des Organismus auf zellulärer Ebene bereitgestellt. Zellen vielzelliger Organismen können in somatische und reproduktive Zellen unterteilt werden.
Somatische Zellen- Dies sind alle Zellen des Körpers, die durch mitotische Teilung entstehen.
Durch die Untersuchung der Chromosomen konnte festgestellt werden, dass die somatischen Zellen des Körpers jeder biologischen Spezies durch eine konstante Anzahl von Chromosomen gekennzeichnet sind. Ein Mensch hat zum Beispiel 46 davon. Man nennt den Chromosomensatz somatischer Zellen diploid(2n) oder doppelt.

Geschlechtszellen, oder Gameten sind spezialisierte Zellen, die zur sexuellen Fortpflanzung dienen.
Gameten enthalten immer halb so viele Chromosomen wie Körperzellen (beim Menschen - 23), daher wird der Chromosomensatz von Keimzellen genannt haploid(n) oder Single. Seine Bildung ist mit der meiotischen Zellteilung verbunden.
Die DNA-Menge in Körperzellen wird mit 2c und in Geschlechtszellen mit 1c bezeichnet. Die genetische Formel somatischer Zellen wird als 2n2c und der Genitalzellen als 1n1c geschrieben.
In den Kernen einiger somatischer Zellen kann die Anzahl der Chromosomen von der Anzahl in somatischen Zellen abweichen. Wenn dieser Unterschied größer als eins, zwei, drei usw. ist, werden solche Zellen als haploide Mengen bezeichnet polyploid(tri-, tetra- bzw. pentaploid). In solchen Zellen laufen Stoffwechselprozesse meist sehr intensiv ab.
Die Anzahl der Chromosomen an sich ist kein artspezifisches Merkmal, da verschiedene Organismen die gleiche Anzahl an Chromosomen haben können, verwandte Organismen jedoch eine unterschiedliche Anzahl. Beispielsweise haben das Malariaplasmodium und der Pferdespulwurm jeweils zwei Chromosomen, während Menschen und Schimpansen 46 bzw. 48 haben.
Menschliche Chromosomen werden in zwei Gruppen eingeteilt: Autosomen und Geschlechtschromosomen (Heterochromosomen). Autosome In menschlichen Körperzellen gibt es 22 Paare, sie sind für Männer und Frauen gleich und Geschlechtschromosomen nur ein Paar, aber dieses bestimmt das Geschlecht des Individuums. Es gibt zwei Arten von Geschlechtschromosomen – X und Y. Die Körperzellen von Frauen tragen zwei X-Chromosomen und die von Männern – X und Y.
Karyotyp- Dies ist eine Reihe von Merkmalen des Chromosomensatzes eines Organismus (die Anzahl der Chromosomen, ihre Form und Größe).
Die bedingte Aufzeichnung eines Karyotyps umfasst die Gesamtzahl der Chromosomen, Geschlechtschromosomen und mögliche Abweichungen im Chromosomensatz. Der Karyotyp eines normalen Mannes wird beispielsweise als 46, XY geschrieben, und der Karyotyp einer normalen Frau ist 46, XX.
Zelllebenszyklus: Interphase und Mitose
Zellen entstehen nicht jedes Mal neu, sie entstehen erst durch die Teilung der Mutterzellen. Nach der Teilung benötigen die Tochterzellen einige Zeit, um Organellen zu bilden und die entsprechende Struktur zu erlangen, die die Erfüllung einer bestimmten Funktion gewährleistet. Dieser Zeitraum wird aufgerufen Reifung.
Als bezeichnet wird die Zeitspanne vom Entstehen einer Zelle infolge der Teilung bis zu ihrer Teilung bzw. ihrem Absterben Lebenszyklus einer Zelle.
In eukaryotischen Zellen ist der Lebenszyklus in zwei Hauptstadien unterteilt: Interphase und Mitose.

Interphase- Dies ist ein Zeitraum im Lebenszyklus, in dem sich die Zelle nicht teilt und normal funktioniert. Die Interphase ist in drei Perioden unterteilt: G 1 -, S- und G 2 -Perioden.
G 1 -Periode(präsynthetisch, postmitotisch) ist eine Phase des Zellwachstums und der Zellentwicklung, in der die aktive Synthese von RNA, Proteinen und anderen Substanzen stattfindet, die für die vollständige Lebenserhaltung der neu gebildeten Zelle notwendig sind. Gegen Ende dieses Zeitraums beginnt die Zelle möglicherweise damit, sich auf die Vervielfältigung ihrer DNA vorzubereiten.
IN S-Periode(synthetisch) Der Prozess der DNA-Replikation selbst findet statt. Der einzige Teil des Chromosoms, der keiner Replikation unterliegt, ist das Zentromer. Die resultierenden DNA-Moleküle divergieren daher nicht vollständig, sondern bleiben darin zusammengehalten, und zu Beginn der Teilung hat das Chromosom ein X-förmiges Aussehen. Die genetische Formel einer Zelle nach der DNA-Verdoppelung lautet 2n4c. Auch in der S-Periode verdoppeln sich die Zentriolen des Zellzentrums.
G 2 -Periode(postsynthetisch, prämitotisch) zeichnet sich durch eine intensive Synthese von RNA, Proteinen und ATP aus, die für den Prozess der Zellteilung sowie die Trennung von Zentriolen, Mitochondrien und Plastiden notwendig sind. Bis zum Ende der Interphase bleiben Chromatin und Nukleolus klar unterscheidbar, die Integrität der Kernhülle wird nicht gestört und die Organellen verändern sich nicht.
Einige Körperzellen können ihre Funktionen ein Leben lang erfüllen (Neuronen unseres Gehirns, Muskelzellen des Herzens), während andere nur für kurze Zeit existieren und dann absterben (Darmepithelzellen, Epidermiszellen). die Haut). Folglich muss der Körper ständig Prozesse der Zellteilung und der Bildung neuer Zellen durchlaufen, die tote Zellen ersetzen würden. Teilungsfähige Zellen nennt man Stengel. Im menschlichen Körper kommen sie im roten Knochenmark, in den tiefen Schichten der Epidermis der Haut und an anderen Stellen vor. Mithilfe dieser Zellen können Sie ein neues Organ wachsen lassen, eine Verjüngung erreichen und auch den Körper klonen. Die Aussichten für die Verwendung von Stammzellen sind absolut klar, die moralischen und ethischen Aspekte dieses Problems werden jedoch immer noch diskutiert, da in den meisten Fällen embryonale Stammzellen verwendet werden, die aus menschlichen Embryonen stammen, die bei einer Abtreibung getötet wurden.
Die Dauer der Interphase in pflanzlichen und tierischen Zellen beträgt durchschnittlich 10–20 Stunden, während die Mitose etwa 1–2 Stunden dauert.
Während aufeinanderfolgender Teilungen in mehrzelligen Organismen werden Tochterzellen immer vielfältiger, da sie Informationen aus einer zunehmenden Anzahl von Genen lesen.
Einige Zellen hören mit der Zeit auf, sich zu teilen, und sterben ab. Dies kann auf die Erfüllung bestimmter Funktionen zurückzuführen sein, wie im Fall der epidermalen Hautzellen und Blutzellen, oder auf eine Schädigung dieser Zellen durch Umweltfaktoren, insbesondere Krankheitserreger. Man nennt es genetisch programmierten Zelltod Apoptose, während Unfalltod - Nekrose.
Mitose ist die Teilung somatischer Zellen. Phasen der Mitose
Mitose- eine Methode zur indirekten Teilung somatischer Zellen.
Während der Mitose durchläuft die Zelle eine Reihe aufeinanderfolgender Phasen, in deren Folge jede Tochterzelle denselben Chromosomensatz wie die Mutterzelle erhält.
Die Mitose wird in vier Hauptphasen unterteilt: Prophase, Metaphase, Anaphase und Telophase. Prophase- das längste Stadium der Mitose, in dem Chromatin kondensiert, wodurch X-förmige Chromosomen sichtbar werden, die aus zwei Chromatiden (Tochterchromosomen) bestehen. In diesem Fall verschwindet der Nukleolus, die Zentriolen divergieren zu den Polen der Zelle und es beginnt sich eine Achromatinspindel (Teilungsspindel) aus Mikrotubuli zu bilden. Am Ende der Prophase zerfällt die Kernmembran in einzelne Vesikel.

IN Metaphase Die Chromosomen sind entlang des Äquators der Zelle mit ihren Zentromeren aufgereiht, an denen die Mikrotubuli der vollständig ausgebildeten Spindel befestigt sind. In diesem Teilungsstadium sind die Chromosomen am stärksten verdichtet und haben eine charakteristische Form, die die Untersuchung des Karyotyps ermöglicht.
IN Anaphase An Zentromeren findet eine schnelle DNA-Replikation statt, wodurch Chromosomen gespalten werden und Chromatiden zu den Polen der Zelle auseinanderlaufen, die durch Mikrotubuli gedehnt werden. Die Verteilung der Chromatiden muss absolut gleich sein, da dieser Prozess die Aufrechterhaltung einer konstanten Anzahl von Chromosomen in den Körperzellen gewährleistet.
Auf der Bühne Telophasen Tochterchromosomen sammeln sich an den Polen, um sie herum bilden sich aus Vesikeln spiralförmige Kernmembranen, und in den neu gebildeten Kernen erscheinen Nukleolen.
Nach der Kernteilung erfolgt die zytoplasmatische Teilung – Zytokinese, Dabei kommt es zu einer mehr oder weniger gleichmäßigen Verteilung aller Organellen der Mutterzelle.
So entstehen durch Mitose aus einer Mutterzelle zwei Tochterzellen, die jeweils eine genetische Kopie der Mutterzelle sind (2n2c).
In kranken, beschädigten, alternden Zellen und spezialisierten Geweben des Körpers kann ein etwas anderer Teilungsprozess stattfinden – Amitose. Amitose sogenannte direkte Teilung eukaryotischer Zellen, bei der es nicht zur Bildung genetisch äquivalenter Zellen kommt, da die Zellbestandteile ungleichmäßig verteilt sind. Es kommt bei Pflanzen im Endosperm und bei Tieren vor – in der Leber, im Knorpel und in der Hornhaut des Auges.
Meiose. Phasen der Meiose
Meiose ist eine Methode der indirekten Teilung primärer Keimzellen (2n2c), die zur Bildung haploider Zellen (1n1c), meist Keimzellen, führt.
Im Gegensatz zur Mitose besteht die Meiose aus zwei aufeinanderfolgenden Zellteilungen, denen jeweils eine Interphase vorausgeht. Der erste Abschnitt der Meiose (Meiose I) wird genannt reduktionistisch, da in diesem Fall die Anzahl der Chromosomen halbiert wird und die zweite Teilung (Meiose II) - gleichwertig, da dabei die Anzahl der Chromosomen erhalten bleibt.

Interphase I verläuft wie die Interphase der Mitose. Meiose I ist in vier Phasen unterteilt: Prophase I, Metaphase I, Anaphase I und Telophase I. B Prophase I Es finden zwei wichtige Prozesse statt: Konjugation und Crossing Over. Konjugation- Dies ist der Prozess der Verschmelzung homologer (gepaarter) Chromosomen über die gesamte Länge. Die bei der Konjugation gebildeten Chromosomenpaare bleiben bis zum Ende der Metaphase I erhalten.
Überqueren- gegenseitiger Austausch homologer Bereiche homologer Chromosomen. Durch das Überkreuzen erhalten die Chromosomen, die der Körper von beiden Elternteilen erhält, neue Genkombinationen, was zur Entstehung genetisch vielfältiger Nachkommen führt. Am Ende der Prophase I verschwindet wie in der Prophase der Mitose der Nukleolus, die Zentriolen divergieren zu den Polen der Zelle und die Kernmembran zerfällt.

IN Metaphase I Chromosomenpaare sind entlang des Äquators der Zelle ausgerichtet und Spindelmikrotubuli sind an ihren Zentromeren befestigt.
IN Anaphase I Ganze homologe Chromosomen, bestehend aus zwei Chromatiden, divergieren zu den Polen.
IN Telophase I Um die Chromosomenhaufen an den Zellpolen bilden sich Kernmembranen und es bilden sich Nukleolen.
Zytokinese I sorgt für die Trennung des Zytoplasmas von Tochterzellen.
Die durch Meiose I entstandenen Tochterzellen (1n2c) sind genetisch heterogen, da ihre Chromosomen, zufällig an den Zellpolen verteilt, unterschiedliche Gene enthalten.
Vergleichende Merkmale von Mitose und Meiose
| Zeichen | Mitose | Meiose | |
| Welche Zellen beginnen sich zu teilen? | Somatisch (2n) | Primäre Keimzellen (2n) | |
| Anzahl der Divisionen | 1 | 2 | |
| Wie viele und welche Art von Zellen entstehen bei der Teilung? | 2 somatisch (2n) | 4 sexuell(n) | |
| Interphase | Vorbereitung der Zelle auf die Teilung, DNA-Verdoppelung | Sehr kurz, es findet keine DNA-Verdoppelung statt | |
| Phasen | Meiose I | Meiose II | |
| Prophase | Es kann zu Chromosomenkondensation, Verschwinden des Nukleolus, Zerfall der Kernmembran, Konjugation und Crossing-over kommen | Chromosomenkondensation, Verschwinden des Nukleolus, Zerfall der Kernmembran | |
| Metaphase | Entlang des Äquators liegen Chromosomenpaare, es entsteht eine Spindel | Chromosomen reihen sich entlang des Äquators auf, es entsteht eine Spindel | |
| Anaphase | Homologe Chromosomen zweier Chromatiden bewegen sich in Richtung der Pole | Chromatiden bewegen sich in Richtung der Pole | |
| Telophase | Chromosomen despiral, neue Kernmembranen und Nukleolen werden gebildet | Chromosomen despiral, neue Kernmembranen und Nukleolen werden gebildet | |
Interphase II sehr kurz, da darin keine DNA-Verdoppelung stattfindet, also keine S-Periode vorhanden ist.
Meiose II ebenfalls in vier Phasen unterteilt: Prophase II, Metaphase II, Anaphase II und Telophase II. IN Prophase II Es finden die gleichen Prozesse wie in der Prophase I statt, mit Ausnahme der Konjugation und des Crossing-Over.
IN Metaphase II Chromosomen befinden sich entlang des Äquators der Zelle.
IN Anaphase II Chromosomen werden an den Zentromeren gespalten und die Chromatiden werden zu den Polen hin gestreckt.
IN Telophase II Kernmembranen und Nukleolen werden um Ansammlungen von Tochterchromosomen herum gebildet.
Nach Zytokinese II Die genetische Formel aller vier Tochterzellen ist 1n1c, aber sie haben alle einen unterschiedlichen Satz von Genen, der das Ergebnis der Überkreuzung und der zufälligen Kombination der Chromosomen der mütterlichen und väterlichen Organismen in den Tochterzellen ist.
Entwicklung von Keimzellen bei Pflanzen und Tieren
Gametogenese(aus dem Griechischen Gamet- Gattin, Gameten- Ehemann und Genesis- Ursprung, Entstehung) ist der Prozess der Bildung reifer Keimzellen.
Da für die sexuelle Fortpflanzung meist zwei Individuen – ein Weibchen und ein Männchen – erforderlich sind, die unterschiedliche Geschlechtszellen – Eier und Spermien – produzieren, müssen die Prozesse der Bildung dieser Gameten unterschiedlich sein.
Die Art des Prozesses hängt in erheblichem Maße davon ab, ob er in einer pflanzlichen oder tierischen Zelle abläuft, da bei Pflanzen während der Gametenbildung nur Mitose auftritt und bei Tieren sowohl Mitose als auch Meiose.
Entwicklung von Keimzellen in Pflanzen. Bei Angiospermen erfolgt die Bildung männlicher und weiblicher Fortpflanzungszellen in verschiedenen Teilen der Blüte – den Staubblättern bzw. Stempeln.
Vor der Bildung männlicher Fortpflanzungszellen - Mikrogametogenese(aus dem Griechischen Mikros- klein) - passiert Mikrosporogenese, also die Bildung von Mikrosporen in den Staubbeuteln der Staubblätter. Dieser Prozess ist mit der meiotischen Teilung der Mutterzelle verbunden, die zur Bildung von vier haploiden Mikrosporen führt. Die Mikrogametogenese ist mit der mitotischen Teilung der Mikrospore verbunden, wodurch aus zwei Zellen ein männlicher Gametophyt entsteht – ein großer vegetativ(siphonogen) und flach generativ. Nach der Teilung wird der männliche Gametophyt mit dichten Membranen bedeckt und bildet ein Pollenkörner. In einigen Fällen teilt sich die generative Zelle sogar während des Prozesses der Pollenreifung und manchmal erst nach der Übertragung auf die Narbe des Stempels mitotisch, um zwei unbewegliche männliche Keimzellen zu bilden – Sperma. Nach der Bestäubung bildet sich aus der vegetativen Zelle ein Pollenschlauch, durch den Spermien zur Befruchtung in den Eierstock des Stempels eindringen.

Als bezeichnet wird die Entwicklung weiblicher Keimzellen in Pflanzen Megagametogenese(aus dem Griechischen Megas- groß). Es kommt im Eierstock des Stempels vor, dem vorangestellt ist Megasporogenese, wodurch aus der im Nucellus liegenden Mutterzelle der Megaspore durch meiotische Teilung vier Megasporen entstehen. Eine der Megasporen teilt sich mitotisch dreimal, wodurch der weibliche Gametophyt entsteht – ein Embryosack mit acht Kernen. Durch die anschließende Trennung des Zytoplasmas der Tochterzellen wird aus einer der resultierenden Zellen eine Eizelle, an deren Seiten die sogenannten Synergiden liegen, am gegenüberliegenden Ende des Embryosacks drei Antipoden gebildet werden und in der Mitte Durch die Verschmelzung zweier haploider Kerne entsteht eine diploide Zentralzelle.
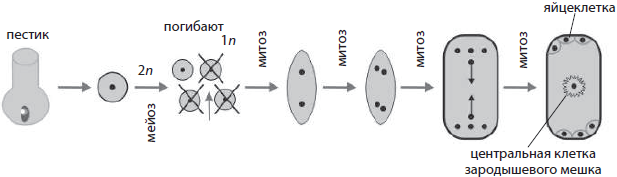
Entwicklung von Keimzellen bei Tieren. Bei Tieren gibt es zwei Prozesse der Keimzellenbildung – Spermatogenese und Oogenese.
Spermatogenese(aus dem Griechischen Sperma, Spermatos- Saatgut und Genesis- Herkunft, Vorkommen) ist der Prozess der Bildung reifer männlicher Keimzellen - Spermien. Beim Menschen kommt es in den Hoden vor und wird in vier Phasen unterteilt: Fortpflanzung, Wachstum, Reifung und Bildung.

IN Brutzeit Urkeimzellen teilen sich mitotisch, was zur Bildung von Diploiden führt Spermatogonie. IN Wachstumsphase Spermatogonien sammeln Nährstoffe im Zytoplasma an, nehmen an Größe zu und verwandeln sich in primäre Spermatozyten, oder Spermatozyten 1. Ordnung. Erst danach treten sie in die Meiose ein ( Reifezeit), wodurch die ersten beiden gebildet werden sekundärer Spermatozyten, oder Spermatozyten 2. Ordnung, und dann - vier haploide Zellen mit immer noch ziemlich viel Zytoplasma - Spermatiden. IN Entstehungszeit Sie verlieren fast ihr gesamtes Zytoplasma und bilden ein Flagellum, das sich in Spermien verwandelt.
Sperma, oder lebhaft, - sehr kleine bewegliche männliche Fortpflanzungszellen mit Kopf, Hals und Schwanz.

IN Kopf, zusätzlich zum Kern, ist Akrosom- ein modifizierter Golgi-Komplex, der für die Auflösung der Eimembranen während der Befruchtung sorgt. IN Gebärmutterhals sind die Zentriolen des Zellzentrums und der Basis Pferdeschwanz bilden Mikrotubuli, die die Spermienbewegung direkt unterstützen. Es enthält auch Mitochondrien, die die Spermien mit ATP-Energie für ihre Bewegung versorgen.
Oogenese(aus dem Griechischen UN- Ei und Genesis- Herkunft, Vorkommen) ist der Prozess der Bildung reifer weiblicher Keimzellen - Eier. Beim Menschen findet es in den Eierstöcken statt und besteht aus drei Phasen: Fortpflanzung, Wachstum und Reifung. Während der intrauterinen Entwicklung treten Fortpflanzungs- und Wachstumsperioden auf, die denen der Spermatogenese ähneln. In diesem Fall entstehen durch Mitose aus primären Keimzellen diploide Zellen. Oogonie, die sich dann in diploide Primärzellen verwandeln Eizellen, oder Eizellen 1. Ordnung. Meiose und anschließende Zytokinese treten auf Reifezeit, zeichnen sich durch eine ungleichmäßige Teilung des Zytoplasmas der Mutterzelle aus, so dass zunächst eine solche erhalten wird sekundäre Eizelle, oder Eizelle 2. Ordnung, Und erster Polkörper, und dann aus der sekundären Eizelle – der Eizelle, die den gesamten Nährstoffvorrat behält, und dem zweiten Polkörper, während der erste Polkörper in zwei Teile geteilt ist. Polkörperchen nehmen überschüssiges genetisches Material auf.
Beim Menschen werden Eier im Abstand von 28 bis 29 Tagen produziert. Der Zyklus, der mit der Reifung und Freisetzung der Eizellen verbunden ist, wird Menstruation genannt.
Ei- eine große weibliche Fortpflanzungszelle, die nicht nur einen haploiden Chromosomensatz, sondern auch einen erheblichen Nährstoffvorrat für die spätere Entwicklung des Embryos trägt.

Das Ei von Säugetieren ist mit vier Membranen bedeckt, die die Wahrscheinlichkeit einer Schädigung durch verschiedene Faktoren verringern. Der Durchmesser des Eies erreicht beim Menschen 150-200 Mikrometer, während er beim Strauß mehrere Zentimeter betragen kann.
Die Zellteilung ist die Grundlage für das Wachstum, die Entwicklung und die Fortpflanzung von Organismen. Die Rolle von Mitose und Meiose
Führt die Zellteilung bei einzelligen Organismen zu einer Zunahme der Individuenzahl, also zur Fortpflanzung, so kann dieser Vorgang bei mehrzelligen Organismen unterschiedliche Bedeutungen haben. Somit ist die Teilung embryonaler Zellen ausgehend von der Zygote die biologische Grundlage der miteinander verbundenen Wachstums- und Entwicklungsprozesse. Ähnliche Veränderungen werden beim Menschen im Jugendalter beobachtet, wenn nicht nur die Zellzahl zunimmt, sondern auch eine qualitative Veränderung im Körper auftritt. Auch die Fortpflanzung vielzelliger Organismen basiert auf der Zellteilung, beispielsweise wird bei der asexuellen Fortpflanzung durch diesen Prozess ein ganzer Teil des Organismus wiederhergestellt, und bei der sexuellen Fortpflanzung werden im Prozess der Gametogenese Geschlechtszellen gebildet, die Anschließend entsteht ein neuer Organismus. Es ist zu beachten, dass die wichtigsten Teilungsmethoden einer eukaryontischen Zelle – Mitose und Meiose – im Lebenszyklus von Organismen unterschiedliche Bedeutungen haben.
Durch die Mitose kommt es zu einer gleichmäßigen Verteilung des Erbmaterials zwischen den Tochterzellen – exakten Kopien der Mutter. Ohne Mitose wäre die Existenz und das Wachstum mehrzelliger Organismen, die sich aus einer einzigen Zelle, der Zygote, entwickeln, unmöglich, da alle Zellen solcher Organismen die gleiche genetische Information enthalten müssen.
Im Zuge der Teilung werden Tochterzellen in Struktur und Funktion immer vielfältiger, was mit der Aktivierung immer neuer Gengruppen in ihnen durch interzelluläre Interaktion einhergeht. Daher ist Mitose für die Entwicklung des Organismus notwendig.
Diese Methode der Zellteilung ist für die Prozesse der ungeschlechtlichen Fortpflanzung und Regeneration (Wiederherstellung) geschädigter Gewebe und Organe notwendig.
Die Meiose wiederum sorgt für die Konstanz des Karyotyps bei der sexuellen Fortpflanzung, da sie den Chromosomensatz vor der sexuellen Fortpflanzung halbiert, der dann durch die Befruchtung wiederhergestellt wird. Darüber hinaus führt die Meiose durch Kreuzung und zufällige Kombination von Chromosomen in Tochterzellen zur Entstehung neuer Kombinationen von Elterngenen. Dadurch erweist sich der Nachwuchs als genetisch vielfältig, was Material für die natürliche Selektion liefert und die materielle Grundlage für die Evolution darstellt. Eine Veränderung der Anzahl, Form und Größe der Chromosomen kann einerseits zum Auftreten verschiedener Abweichungen in der Entwicklung des Organismus und sogar zu dessen Tod führen, andererseits kann sie zum Auftreten von Individuen führen besser an die Umgebung angepasst.
Somit ist die Zelle die Einheit für Wachstum, Entwicklung und Fortpflanzung von Organismen.
§ 2. Hauptbestandteile einer eukaryotischen Zelle
Eukaryontische Zellen (Abb. 8 und 9) sind viel komplexer organisiert als prokaryontische Zellen. Sie sind sehr unterschiedlich in der Größe (von mehreren Mikrometern bis zu mehreren Zentimetern), in der Form und in den Strukturmerkmalen (Abb. 10).
Reis. 8. Struktur einer eukaryotischen Zelle. Verallgemeinertes Schema
Reis. 9. Zellstruktur gemäß Elektronenmikroskopie
Reis. 10. Verschiedene eukaryontische Zellen: 1 – Epithelzellen; 2 – Blut (e – Erythroitis, / – Leukitis); 3 – Knorpel; 4 – Knochen; 5 – glatte Muskulatur; 6 – Bindegewebe; 7 – Nervenzellen; 8 – quergestreifte Muskelfaser
Die allgemeine Organisation und das Vorhandensein grundlegender Komponenten sind jedoch in allen eukaryotischen Zellen gleich (Abb. 11).
Reis. 11. Eukaryontische Zelle (Diagramm)
Plasmalemma (äußere Zellmembran). Die Basis der Plasmamembran ist, wie auch anderer Membranen in Zellen (zum Beispiel Mitochondrien, Plastiden etc.), eine Lipidschicht, die aus zwei Molekülreihen besteht (Abb. 12). Da Lipidmoleküle polar sind (ein Pol ist hydrophil, d. h. von Wasser angezogen, und der andere ist hydrophob, d. h. von Wasser abgestoßen), sind sie in einer bestimmten Reihenfolge angeordnet. Die hydrophilen Enden der Moleküle einer Schicht sind in Richtung der wässrigen Umgebung gerichtet – in das Zytoplasma der Zelle, und die andere Schicht – von der Zelle nach außen – in Richtung der interzellulären Substanz (bei mehrzelligen Organismen) oder der wässrigen Umgebung (bei einzelligen Organismen). ).
Reis. 12. Aufbau der Zellmembran nach dem Fluidmosaikmodell. Proteine und Glykoproteine sind in eine Doppelschicht aus Lipidmolekülen eingetaucht, wobei ihre hydrophilen Enden (Kreise) nach außen zeigen und ihre hydrophoben Enden (Wellenlinien) tief in die Membran hinein zeigen
Proteinmoleküle sind mosaikartig in eine bimolekulare Lipidschicht eingebettet. An der Außenseite der tierischen Zelle lagern sich Polysaccharidmoleküle an die Lipide und Proteinmoleküle des Plasmalemmas an und bilden Glykolipide und Glykoproteine.
Dieses Aggregat bildet eine Schicht Glykokalyx. Mit ihm verbunden Rezeptorfunktion Plasmamembranen (siehe unten); Es kann auch verschiedene von der Zelle verwendete Substanzen ansammeln. Darüber hinaus erhöht die Glykokalyx die mechanische Stabilität des Plasmalemmas.
Auch in den Zellen von Pflanzen und Pilzen gibt es eine Zellwand, die eine unterstützende und schützende Rolle spielt. Bei Pflanzen besteht es aus Zellulose, bei Pilzen aus Chitin.
Die äußere Zellmembran erfüllt eine Reihe von Funktionen, darunter:
♦ mechanisch(unterstützend, formbildend);
♦ Barriere-Transport(selektive Durchlässigkeit für verschiedene Substanzen: Eintritt notwendiger Substanzen in die Zelle und Entfernung unnötiger und schädlicher Substanzen);
♦ Rezeptor(Bestimmung verschiedener Chemikalien in unmittelbarer Nähe der Zelle; Wahrnehmung von Signalen in Form von Hormonen; Erkennung „fremder“ Proteine durch Zellen des Immunsystems usw.).
Der Stoffaustausch zwischen Zelle und Umwelt erfolgt auf unterschiedliche Weise – passiv und aktiv.
Durch spezielle Poren dringen Wassermoleküle und verschiedene Ionen passiv (durch Diffusion, Osmose) ein, ohne dass die Zelle Energie verbraucht passiver Transport. Makromoleküle wie Proteine, Polysaccharide und sogar ganze Zellen werden durch versorgt Phagozytose Und Pinozytose mit Energieverbrauch – aktiven Transport.
Durch die Phagozytose werden ganze Zellen oder große Partikel verschlungen (denken Sie zum Beispiel an die Ernährung in Amöben oder die Phagozytose durch schützende Blutzellen von Bakterien). Bei der Pinozytose werden kleine Partikel oder Tröpfchen einer flüssigen Substanz absorbiert. Beiden Prozessen ist gemeinsam, dass die aufgenommenen Stoffe von einer einstülpenden Außenmembran umgeben werden und so eine Vakuole bilden, die dann tief in das Zytoplasma der Zelle vordringt.
Exozytose ist ein Prozess (der auch aktiver Transport ist), der der Phagozytose und Pinozytose entgegengesetzt ist (Abb. 13). Mit seiner Hilfe können unverdaute Nahrungsreste in Protozoen oder in der sekretorischen Zelle gebildete biologisch aktive Substanzen entfernt werden.
Zytoplasma. Zytoplasma ist der durch das Plasmalemma begrenzte Zellinhalt mit Ausnahme des Zellkerns. Es beinhaltet Grundsubstanz (Hyaloplasma), Organellen Und Einschlüsse.
Hyaloplasma- eine viskose Flüssigkeit, die sich in einem der beiden Zustände befinden kann Zola(flüssig), oder Gel(geleeartig).
Bei Bedarf ist das Zytoplasma in der Lage, reversibel von einem Zustand in einen anderen überzugehen. Beispielsweise kommt es während der Amöbenbewegung (denken Sie an den Abschnitt „Protozoen“ aus dem Zoologiekurs) während der Bildung eines Pseudopoden zu schnellen Übergängen des Zytoplasmas vom Gel zum Sol und umgekehrt. Dies ist auf das Vorhandensein einer großen Anzahl filamentöser Proteinmoleküle im Zytoplasma zurückzuführen Aktina. Wenn sie sich zu einem dreidimensionalen Netzwerk verbinden, befindet sich das Zytoplasma in einem Gelzustand, und wenn das Netzwerk zerfällt, befindet es sich in einem Solzustand.
Das Hyaloplasma enthält verschiedene Substanzen – Enzyme, Proteine, Kohlenhydrate, Fette und andere, organische und mineralische. Hier finden verschiedene chemische Prozesse statt – der Abbau von Stoffen, deren Synthese und Modifikationen (Veränderungen).
Organoide. Dabei handelt es sich um dauerhafte Bestandteile einer Zelle mit einer bestimmten Struktur und Funktion, die sich in ihrem Zytoplasma befinden. Im Folgenden werden wir darüber sprechen Allzweck-Organoide, allen Zelltypen aller Eukaryoten inhärent. Sie sind mit der Sicherstellung der lebenswichtigen Funktionen des letzteren verbunden. Organoide für besondere Zwecke kommt nur in Zellen eines bestimmten (hochspezialisierten) Typs vor – zum Beispiel Myofibrillen in Muskelzellen.
Allzweckorganellen haben die gleiche Struktur, unabhängig davon, zu welchen Zellen und Organismen sie gehören. Aber unter ihnen gibt es Gruppen mit Membran (Endoplasmatisches Retikulum, Golgi-Apparat, Mitochondrien, Plastiden, Lysosomen, Vakuolen), sowie Nichtmembran ( Ribosomen, Zellzentrum) Struktur.
Endoplasmatisches Retikulum (ER). Das EPS besteht aus Membranen und ist ein komplexes verzweigtes System aus Tubuli und Zisternen, das das gesamte Zytoplasma der Zelle durchdringt (Abb. 14). Es gibt zwei Arten von EPS: rauh Und glatt. Ribosomen sind an den rauen Membranen befestigt (von der zytoplasmatischen Seite), während sie auf den glatten Membranen nicht vorhanden sind.
Reis. 14. Endoplasmatisches Retikulum
Das endoplasmatische Retikulum erfüllt in einer eukaryotischen Zelle eine Reihe wichtiger Funktionen:
♦ abgrenzen(Aufteilung des Innenvolumens der Zelle in verschiedene Reaktionsräume);
♦ Beteiligung an der Synthese organischer Stoffe(Ribosomen befinden sich auf den Membranen des rauen ER und Enzymkomplexe auf den glatten Membranen, die die Synthese von Lipiden, Kohlenhydraten usw. gewährleisten);
♦ Beteiligung an der Bildung von Elementen des Golgi-Apparats, Lysosomen;
♦ Transport von Stoffen.
Golgi-Apparat. Der Golgi-Apparat (AG) ist ein System Panzer(flache Vakuolen) und Blasen(Vesikel), die sich in unmittelbarer Nähe des Zellkerns befinden und aufgrund der EPS durch die Trennung seiner kleinen Fragmente entstehen (Abb. 15). Wenn diese Fragmente verschmelzen, entstehen neue Zisternen des Golgi-Apparats, während verschiedene Substanzen aus dem EPS transportiert werden, die am Aufbau komplexer organischer Verbindungen (Proteine + Kohlenhydrate, Proteine + Lipide usw.) beteiligt sind, die mit dem entfernt werden Hilfe von AG außerhalb der Zelle. Diese biologisch aktiven Substanzen werden entweder aus der Zelle entfernt (unter Verwendung sekretorischer Vakuolen durch Exozytose) oder sind Teil von Lysosomen (siehe unten), die von AG gebildet werden.
Reis. 15. Golgi-Apparat:
Der Golgi-Apparat erfüllt folgende Funktionen:
♦ Synthese von der Zelle produzierte biologisch aktive Substanzen;
♦ Sekretion (Entfernung aus der Zelle) verschiedener Substanzen(Hormone, Enzyme, Stoffe, aus denen die Zellwand aufgebaut ist usw.);
♦ Beteiligung an der Bildung von Lysosomen.
Mitochondrien. Alle Arten eukaryotischer Zellen verfügen über Mitochondrien (Abb. 16). Sie sehen entweder wie runde Körper oder Stäbe aus, seltener wie Fäden. Ihre Größe reicht von 1 bis 7 Mikrometer. Die Anzahl der Mitochondrien in einer Zelle liegt zwischen mehreren Hundert und Zehntausenden (bei großen Protozoen).
Reis. 16. Mitochondrien. Oben - Mitochondrien (a) in den Harnkanälen, sichtbar im Lichtmikroskop. Unten ist ein dreidimensionales Modell der mitochondrialen Organisation: 1 – Cristae; 2 – äußere Membran; 3 – innere Membran; 4 – Matrix
Das Mitochondrium besteht aus zwei Membranen – extern Und intern, zwischen denen liegt Zwischenmembranraum. Die innere Membran bildet viele Einstülpungen – Cristae, die entweder Platten oder Röhren sind. Diese Organisation stellt einen riesigen Bereich der inneren Membran bereit. Es enthält Enzyme, die für die Umwandlung der in organischen Substanzen (Kohlenhydrate, Lipide) enthaltenen Energie in ATP-Energie sorgen, die für das Leben der Zelle notwendig ist. Daher besteht die Funktion der Mitochondrien darin, daran teilzunehmen Energie zelluläre Prozesse. Deshalb sind beispielsweise in Muskelzellen, die viel Arbeit verrichten, zahlreiche Mitochondrien vorhanden.
Plastiden. In Pflanzenzellen gibt es spezielle Organellen – Plastiden, die oft eine spindelförmige oder abgerundete Form haben, manchmal komplexer. Es gibt drei Arten von Plastiden – Chloroplasten (Abb. 17), Chromoplasten und Leukoplasten.
Chloroplasten unterscheiden sich in der grünen Farbe, was auf das Pigment zurückzuführen ist - Chlorophyll, Sicherstellung des Prozesses Photosynthese, also die Synthese organischer Stoffe aus Wasser (H 2 O) und Kohlendioxid (CO 2) unter Nutzung der Energie des Sonnenlichts. Chloroplasten kommen hauptsächlich in Blattzellen (bei höheren Pflanzen) vor. Sie bestehen aus zwei parallel zueinander angeordneten Membranen, die den Inhalt von Chloroplasten umgeben - Stroma. Die innere Membran bildet zahlreiche abgeflachte Säcke - Thylakoide, die gestapelt sind (wie ein Stapel Münzen) – Körner – und liegen im Stroma. Es sind die Thylakoide, die Chlorophyll enthalten.
Chromoplasten bestimmen die gelbe, orange und rote Farbe vieler Blüten und Früchte, in deren Zellen sie in großen Mengen vorkommen. Die Hauptpigmente in ihrer Zusammensetzung sind Carotine. Der funktionelle Zweck von Chromoplasten besteht darin, Tiere mit Farbe anzulocken und so die Bestäubung von Blüten und die Verbreitung von Samen sicherzustellen.
Reis. 17. Plastiden: a – Chloroplasten in den Zellen des Elodea-Blattes, sichtbar im Lichtmikroskop; b – Diagramm der inneren Struktur des Chloroplasten mit Grana, das sind Stapel flacher Säcke, die senkrecht zur Oberfläche des Chloroplasten angeordnet sind; c – ein detaillierteres Diagramm, das Anastomosierungsschläuche zeigt, die einzelne Ventilatorkammern verbinden
Leukoplasten sind farblose Plastiden, die in den Zellen unterirdischer Pflanzenteile (zum Beispiel in Kartoffelknollen), Samen und dem Kern von Stängeln enthalten sind. In Leukoplasten wird Stärke hauptsächlich aus Glukose gebildet und in den Speicherorganen von Pflanzen angereichert.
Plastiden einer Art können sich in eine andere verwandeln. Wenn sich beispielsweise im Herbst die Blätter verfärben, verwandeln sich Chloroplasten in Chromoplasten.
Lysosomen. Diese Organellen sehen aus wie Vesikel, die von einer Membran mit einem Durchmesser von bis zu 2 Mikrometern umgeben sind. Sie enthalten mehrere Dutzend Enzyme, die Proteine, Nukleinsäuren, Polysaccharide und Lipide abbauen. Die Funktion von Lysosomen besteht darin, an den Prozessen des intrazellulären Abbaus komplexer organischer Verbindungen (z. B. Nährstoffe oder Substanzen „verbrauchter“ Zellbestandteile) teilzunehmen. Lysosomen verschmelzen mit phagozytischen (oder pinozytischen) Vakuolen und bilden eine Verdauungsvakuole.
Die Bildung von Lysosomen erfolgt durch Knospung aus den Zisternen des Golgi-Apparats.
Ribosomen. Ribosomen (Abb. 18) kommen in den Zellen sowohl von Eukaryoten als auch von Prokaryoten vor, da sie eine wichtige Funktion darin erfüllen Proteinbiosynthese(siehe Kapitel 5). Jede Zelle enthält Zehntausende, Hunderttausende (bis zu mehreren Millionen) dieser kleinen runden Organellen.
Reis. 18. Schema der Struktur eines Ribosoms, das auf der Membran des endoplasmatischen Retikulums sitzt: 1 – kleine Untereinheit; 2 – tRNA; 3 – Aminoacyl-tRNA; 4 – Aminosäure; 5 – große Untereinheit; 6 – Membran des endoplasmatischen Retikulums; 7 – synthetisierte Polypeptidkette
Das Ribosom besteht aus zwei ungleichen Untereinheiten (Teilen). Sie werden getrennt gebildet und verbinden sich während der Synthese eines Proteinmoleküls „umfassend“ mit der Boten-RNA. Ribosomen enthalten verschiedene Proteine und ribosomale RNAs.
Zelluläre Einschlüsse. Als solche bezeichnet man die in der Zelle nicht permanenten Bestandteile, die in der Hauptsubstanz des Zytoplasmas in Form von Körnern, Körnern oder Tröpfchen vorliegen. Einschlüsse können von einer Membran umgeben sein oder auch nicht.
Funktionell gibt es drei Arten von Einschlüssen: Nährstoffe speichern(Stärke, Glykogen, Fette, Proteine), sekretorische Einschlüsse(Substanzen, die für Drüsenzellen charakteristisch sind und von ihnen produziert werden - Hormone der endokrinen Drüsen usw.) und Einbeziehung von Sonderzwecken(in hochspezialisierten Zellen, zum Beispiel Hämoglobin in roten Blutkörperchen).
§ 3. Organisation des Zellkerns. Chromosomen
Der Zellkern (siehe Abb. 8 und 9) ist für das Leben der Zelle von größter Bedeutung, da er als Aufbewahrungsort für die in den Chromosomen enthaltenen Erbinformationen dient (siehe unten).
Der Kern wird von einer Kernhülle begrenzt, die seinen Inhalt trennt (Karyoplasma) aus dem Zytoplasma. Die Hülle besteht aus zwei Membranen, die durch einen Spalt getrennt sind. Beide sind von zahlreichen Poren durchzogen, wodurch der Stoffaustausch zwischen Zellkern und Zytoplasma möglich ist. Im Zellkern der meisten Eukaryoten gibt es 1 bis 7 Nukleolen. Mit ihnen sind die Prozesse der RNA- und tRNA-Synthese verbunden.
Hauptkernkomponenten – Chromosomen, gebildet aus einem DNA-Molekül und verschiedenen Proteinen. Im Lichtmikroskop sind sie nur während der Zellteilung deutlich sichtbar (Mitose, Meiose). In einer sich nicht teilenden Zelle sehen die Chromosomen wie lange, dünne Fäden aus, die über das gesamte Zellkernvolumen verteilt sind.
Bei der Zellteilung bilden Chromosomenstränge dichte Spiralen, wodurch sie (unter einem herkömmlichen Mikroskop) in Form von Stäbchen, „Haarnadeln“, sichtbar werden. Der gesamte Umfang der genetischen Information ist auf die Chromosomen des Zellkerns verteilt. Bei ihrer Untersuchung wurden folgende Muster identifiziert:
♦ In den Kernen somatischer Zellen (d. h. Körperzellen, nicht-sexuell) enthalten alle Individuen derselben Art die gleiche Anzahl an Chromosomen, aus denen sie bestehen Chromosomensatz(Abb. 19);
Reis. 19. Chromosomen verschiedener Pflanzen- und Tierarten, dargestellt im gleichen Maßstab: 1,2 – Amöbe; 3,4 – Kieselalgen; 5–8, 18,19 – Grünalgen; 9 – Fliegenpilz; 10 – Linde; 11–12 – Drosophila; 13 – Lachs; 14 – Skerda (Familie der Korbblütler); 15 – Pflanze aus der Familie der Schilddrüsen; 16 – Corydalis-Schmetterling; 17 – Insekt aus der Familie der Heuschrecken; 20 – Wasserläuferkäfer; 21 – Blumenkäfer; 22 – Amphibien-Ambystoma; 23 – Aloe (Liliengewächs)
♦ Jede Art zeichnet sich durch einen eigenen Chromosomensatz entsprechend ihrer Anzahl aus (zum Beispiel hat ein Mensch 46 Chromosomen, eine Drosophila-Fliege hat 8, ein Spulwurm hat 4, ein Flusskrebs hat 196, ein Pferd hat 66 und Mais hat 104);
♦ Chromosomen in den Kernen somatischer Zellen können in sogenannten Paaren gruppiert werden homologe Chromosomen basierend auf ihrer Ähnlichkeit (in Struktur und Funktion);
♦ In den Kernen von Keimzellen (Gameten) enthält jedes Paar homologer Chromosomen nur eines, d. h. der gesamte Chromosomensatz ist halb so groß wie der von Körperzellen.
♦ wird ein einzelner Chromosomensatz in Keimzellen genannt haploid und wird mit dem Buchstaben n bezeichnet, und in somatischer Form - diploid(2n).
Aus dem oben Gesagten geht klar hervor, dass jedes Paar homologer Chromosomen durch die Vereinigung väterlicher und mütterlicher Chromosomen während der Befruchtung entsteht, d. h. durch die Verschmelzung von Geschlechtszellen (Gameten). Umgekehrt gelangt bei der Bildung von Keimzellen von jedem homologen Chromosomenpaar nur eines in die Keimzelle.
Chromosomen anders Homologe Paare unterscheiden sich in Größe und Form (Abb. 20 und 21).
Reis. 20. Struktur und Arten der Chromosomen: a – Aussehen 1 – Zentromer; 2 – kurze Schulter; 3 – lange Schulter); innere Struktur desselben Chromosoms (1 – Zentromer; – DNA-Moleküle); c – Chromosomentypen (1 – einarmig; verschiedenarmig; 3 – gleicharmig: X – Arm, V – Zentromer)
Reis. 21. Ein Chromosom besteht aus DNA und Proteinen. Das DNA-Molekül wird repliziert. Zwei identische DNA-Doppelhelices bleiben im Zentromerbereich verbunden. Diese Kopien werden später während der Zellteilung zu separaten Chromosomen
Im Körper gibt es Chromosomen die primäre Verengung (Zentromer genannt), an dem die Fäden befestigt sind Spaltspindeln. Es teilt das Chromosom in zwei Teile Schulter Chromosomen können gleicharmig, ungleicharmig oder einarmig sein.
Kapitel 5. Stoffwechsel
§ 1. Stoffwechsel als Einheit von Assimilation und Dissimilation
Alle Zellen und lebenden Organismen sind offene Systeme, das heißt, sie stehen in einem ständigen Austausch von Energien und Stoffen mit der Umwelt. In der unbelebten Natur gibt es offene Systeme, deren Existenz sich jedoch qualitativ von der lebender Organismen unterscheidet. Betrachten Sie dieses Beispiel: Ein brennendes Stück nativen Schwefel steht im Austausch mit der Umwelt. Bei der Verbrennung wird O 2 absorbiert und SO 2 sowie Energie (in Form von Wärme) freigesetzt. Allerdings wird in diesem Fall ein Stück Schwefel als physischer Körper zerstört und verliert seine Primärstruktur.
Für lebende Organismen erweist sich der Austausch mit der Umwelt als Voraussetzung für die Erhaltung und Aufrechterhaltung ihrer strukturellen Organisation durch Selbsterneuerung aller Stoffe und Bestandteile, aus denen sie bestehen.
Stoffwechsel (Metabolismus) ist eine Reihe von Prozessen, die in lebenden Organismen ablaufen (Verbrauch, Umwandlung, Ansammlung und Freisetzung von Stoffen und Energie), die ihre lebenswichtige Aktivität, Entwicklung, Wachstum und Fortpflanzung sicherstellen. Während des Stoffwechselprozesses werden die Moleküle, aus denen die Zellen bestehen, abgebaut und synthetisiert; Erneuerung der Zellstrukturen und der Interzellularsubstanz.
Der Stoffwechsel basiert auf miteinander verbundenen Prozessen Assimilation(Anabolismus) und Dissimilation(Katabolismus). Bei der Assimilation (plastischer Austausch) werden aus einfachen Stoffen komplexe Stoffe synthetisiert. Dadurch entstehen in der Zelle alle organischen Substanzen, die für den Aufbau ihrer Strukturbestandteile, Enzymsysteme etc. notwendig sind. Die Assimilation erfolgt immer unter Energieaufwand.
Bei der Dissimilation (Energiestoffwechsel) werden komplexe organische Stoffe in einfachere oder anorganische Stoffe zerlegt. Dabei wird Energie freigesetzt, die von der Zelle für die Durchführung verschiedener Prozesse aufgewendet wird, die ihre lebenswichtige Aktivität sicherstellen (Synthese und Transport von Stoffen, mechanische Arbeit usw.).
Alle lebenden Organismen können in zwei Gruppen eingeteilt werden: Autotrophe Und Heterotrophe, die sich in den Energiequellen und notwendigen Substanzen unterscheiden, um ihre lebenswichtigen Funktionen sicherzustellen.
Autotrophe– Organismen, die mithilfe der Energie des Sonnenlichts organische Verbindungen aus anorganischen Substanzen synthetisieren (z Phototrophen– Pflanzen, Cyanobakterien) oder Energie, die aus der Oxidation mineralischer (anorganischer) Stoffe (wie z. B Chemotrophe– Schwefelbakterien, Eisenbakterien usw.). Dadurch sind sie in der Lage, die für ihre Lebenstätigkeit notwendigen Stoffe selbstständig herzustellen.
§ 2. Dissimilation in anaeroben und aeroben Organismen
Organismen können je nach Art der Dissimilation in zwei Gruppen eingeteilt werden: Aerobier Und Anaerobier. Aerobes (aus dem Griechischen. Dämon– Luft) benötigen zum Leben freien Sauerstoff. In Anaerobiern (griech. ac– negatives Teilchen) ist nicht notwendig. In ihnen erfolgt die Dissimilation durch Fermentation – sauerstofffreier, enzymatischer Abbau organischer Stoffe unter Bildung einfacherer organischer Stoffe und Freisetzung von Energie. Zum Beispiel:
♦ Milchsäuregärung:
C 6 H 12 O 6 + 2H 3 PO 4 + 2ADP → 2PH + 2ATP + 2H 2 O;
♦ alkoholische Gärung:
C 6 H 12 O 6 + 2PH + 2ADP → 2C 2 H 5 OH + 2CO 2 + 2ATP + 2H 2 O.
Die bei der Gärung entstehenden Stoffe sind organisch und enthalten daher noch viel Energie.
Reis. 22. Beziehungen zwischen Assimilation und Dissimilation in autotrophen und heterotrophen Organismen
Bei aeroben Organismen kommt es beim Atmungsprozess in den Mitochondrien zu einem vollständigen Abbau organischer Stoffe (unter Verwendung von O 2) zu den energiearmen Endprodukten CO 2 und H 2 O und es wird eine deutlich größere Energiemenge freigesetzt:
C 6 H 12 0 6 (Glukose) + 0 2 > 6C0 2 + 6H 2 0 + Energie (aufgrund dessen werden 38 ATP-Moleküle synthetisiert).
Betrachten wir in Form verallgemeinerter Schemata den Stoffwechsel autotropher und heterotropher aerober Organismen (Abb. 22).
Assimilation. Seine wichtigsten Prozesse sind Photosynthese Und Proteinbiosynthese.
Der Austausch von Stoffen, die in die Zelle gelangen oder von dieser nach außen abgegeben werden, sowie der Austausch verschiedener Signale mit der Mikro- und Makroumgebung erfolgt über die äußere Membran der Zelle. Bekanntlich ist die Zellmembran eine Lipiddoppelschicht, in die verschiedene Proteinmoleküle eingebettet sind, die als spezialisierte Rezeptoren, Ionenkanäle, Geräte zum aktiven Transport oder Abtransport verschiedener Chemikalien, interzelluläre Kontakte usw. fungieren. In gesunden eukaryotischen Zellen sind Phospholipide verteilt Die Membran ist asymmetrisch: Die äußere Oberfläche besteht aus Sphingomyelin und Phosphatidylcholin, die innere Oberfläche aus Phosphatidylserin und Phosphatidylethanolamin. Die Aufrechterhaltung einer solchen Asymmetrie erfordert Energieaufwand. Im Falle einer Zellschädigung, einer Infektion oder eines Energiemangels reichert sich daher die äußere Oberfläche der Membran mit für sie ungewöhnlichen Phospholipiden an, was für andere Zellen und Enzyme zu einem Signal für eine Zellschädigung mit entsprechender Reaktion darauf wird. Die wichtigste Rolle spielt die lösliche Form der Phospholipase A2, die Arachidonsäure abbaut und aus den oben genannten Phospholipiden Lysoformen erzeugt. Arachidonsäure ist das begrenzende Glied für die Bildung von Entzündungsmediatoren wie Eicosanoiden, und Schutzmoleküle – Pentraxine (C-reaktives Protein (CRP), Vorläufer von Amyloidproteinen) – werden an Lysoformen in der Membran gebunden, woraufhin das Komplement aktiviert wird System entlang des klassischen Weges und der Zellzerstörung.
Die Struktur der Membran trägt dazu bei, die Eigenschaften der inneren Umgebung der Zelle und ihre Unterschiede zur äußeren Umgebung zu bewahren. Dies wird durch die selektive Permeabilität der Zellmembran und das Vorhandensein aktiver Transportmechanismen in ihr gewährleistet. Ihre Störung durch direkte Schädigung, beispielsweise durch Tetrodotoxin, Ouabain, Tetraethylammonium, oder bei unzureichender Energieversorgung der entsprechenden „Pumpen“ führt zu Störungen der Elektrolytzusammensetzung der Zelle, Veränderungen ihres Stoffwechsels, Störungen spezifischer Funktionen - Kontraktion, Weiterleitung von Erregungsimpulsen usw. Eine Störung der zellulären Ionenkanäle (Kalzium, Natrium, Kalium und Chlorid) beim Menschen kann auch genetisch durch Mutation der Gene bedingt sein, die für die Struktur dieser Kanäle verantwortlich sind. Sogenannte Kanalopathien verursachen erbliche Erkrankungen des Nerven-, Muskel- und Verdauungssystems. Übermäßiger Wassereintritt in die Zelle kann zu deren Bruch – Zytolyse – aufgrund einer Perforation der Membran bei Komplementaktivierung oder einem Angriff durch zytotoxische Lymphozyten und natürliche Killerzellen führen.
In die Zellmembran sind viele Rezeptoren eingebaut – Strukturen, die in Kombination mit den entsprechenden spezifischen Signalmolekülen (Liganden) ein Signal in die Zelle übertragen. Dies geschieht durch verschiedene Regulierungskaskaden, die aus enzymatisch aktiven Molekülen bestehen, die nacheinander aktiviert werden und letztendlich zur Umsetzung verschiedener zellulärer Programme wie Wachstum und Proliferation, Differenzierung, Motilität, Alterung und Zelltod beitragen. Es gibt zahlreiche Regulierungskaskaden, deren Anzahl jedoch noch nicht vollständig geklärt ist. Das System aus Rezeptoren und damit verbundenen Regulationskaskaden existiert auch innerhalb der Zelle; Sie bilden ein spezifisches regulatorisches Netzwerk mit Konzentrations-, Verteilungs- und Auswahlpunkten für den weiteren Signalweg in Abhängigkeit vom Funktionszustand der Zelle, ihrem Entwicklungsstadium und der gleichzeitigen Wirkung von Signalen anderer Rezeptoren. Das Ergebnis kann eine Hemmung oder Verstärkung des Signals sein, wodurch es auf einen anderen regulatorischen Weg gelenkt wird. Sowohl der Rezeptorapparat als auch Signaltransduktionswege über Regulationskaskaden, beispielsweise zum Zellkern, können durch einen genetischen Defekt, der als angeborener Defekt auf der Ebene des Organismus auftritt, oder durch eine somatische Mutation in einem bestimmten Zelltyp gestört sein. Diese Mechanismen können durch Infektionserreger und Toxine beschädigt werden und verändern sich auch im Laufe des Alterungsprozesses. Das letzte Stadium davon kann eine Störung der Funktionen der Zelle, der Prozesse ihrer Proliferation und Differenzierung sein.
Auf der Oberfläche von Zellen befinden sich außerdem Moleküle, die eine wichtige Rolle bei den Prozessen der interzellulären Interaktion spielen. Dazu können Zelladhäsionsproteine, Histokompatibilitätsantigene, gewebespezifische, differenzierende Antigene usw. gehören. Veränderungen in der Zusammensetzung dieser Moleküle führen zu Störungen interzellulärer Interaktionen und können die Aktivierung geeigneter Mechanismen zur Eliminierung solcher Zellen bewirken, da sie eine Rolle spielen stellen eine gewisse Gefahr für die Integrität des Körpers dar, da sie als Reservoir für Infektionen, insbesondere virale, oder als potenzielle Auslöser des Tumorwachstums dienen.
Verletzung der Energieversorgung der Zelle
Die Energiequelle in der Zelle ist Nahrung, nach deren Abbau Energie in Endstoffe freigesetzt wird. Der Hauptort der Energieproduktion sind Mitochondrien, in denen Stoffe mit Hilfe von Enzymen der Atmungskette oxidiert werden. Die Oxidation ist der Hauptenergielieferant, da bei der Glykolyse im Vergleich zur Oxidation nicht mehr als 5 % der Energie aus der gleichen Menge an Oxidationssubstraten (Glukose) freigesetzt werden. Etwa 60 % der bei der Oxidation freigesetzten Energie werden durch oxidative Phosphorylierung in hochenergetischen Phosphaten (ATP, Kreatinphosphat) gespeichert, der Rest wird als Wärme abgegeben. Anschließend werden hochenergetische Phosphate von der Zelle für Prozesse wie Pumpenbetrieb, Synthese, Teilung, Bewegung, Sekretion usw. verwendet. Es gibt drei Mechanismen, deren Beschädigung zu einer Störung der Energieversorgung der Zelle führen kann: Der erste ist der Mechanismus der Synthese von Enzymen des Energiestoffwechsels, der zweite ist der Mechanismus der oxidativen Phosphorylierung, der dritte ist der Mechanismus der Energienutzung.
Eine Störung des Elektronentransports in der mitochondrialen Atmungskette oder eine Entkopplung der ADP-Oxidation und -Phosphorylierung mit Verlust des Protonenpotentials, der treibenden Kraft für die ATP-Erzeugung, führt zu einer Schwächung der oxidativen Phosphorylierung in einer Weise, dass der größte Teil der Energie als Wärme verloren geht die Zahl der hochenergetischen Verbindungen nimmt ab. Die Entkopplung von Oxidation und Phosphorylierung unter dem Einfluss von Adrenalin nutzen die Zellen homöothermer Organismen, um die Wärmeproduktion zu steigern und gleichzeitig die Körpertemperatur bei Abkühlung konstant zu halten bzw. bei Fieber zu erhöhen. Bei der Thyreotoxikose werden erhebliche Veränderungen der Mitochondrienstruktur und des Energiestoffwechsels beobachtet. Diese Veränderungen sind zunächst reversibel, aber ab einem bestimmten Punkt werden sie irreversibel: Mitochondrien fragmentieren, zerfallen oder schwellen an, verlieren Kristalle, verwandeln sich in Vakuolen und reichern schließlich Substanzen wie Hyalin, Ferritin, Kalzium und Lipofuszin an. Bei Patienten mit Skorbut verschmelzen Mitochondrien zu Chondrisphären, möglicherweise aufgrund einer Membranschädigung durch Peroxidverbindungen. Unter dem Einfluss ionisierender Strahlung kommt es bei der Umwandlung einer normalen Zelle in eine bösartige Zelle zu erheblichen Schäden an Mitochondrien.
Mitochondrien sind ein leistungsfähiges Depot für Kalziumionen, dessen Konzentration um mehrere Größenordnungen höher ist als die im Zytoplasma. Wenn Mitochondrien geschädigt werden, dringt Kalzium in das Zytoplasma ein und führt zur Aktivierung von Proteinasen mit Schäden an intrazellulären Strukturen und Störungen der Funktionen der entsprechenden Zelle, beispielsweise zu Kalziumkontrakturen oder sogar zum „Kalziumtod“ in Neuronen. Durch die Störung der Funktionsfähigkeit der Mitochondrien nimmt die Bildung freier Radikalperoxidverbindungen stark zu, die eine sehr hohe Reaktivität aufweisen und daher wichtige Bestandteile der Zelle – Nukleinsäuren, Proteine und Lipide – schädigen. Dieses Phänomen wird beim sogenannten oxidativen Stress beobachtet und kann negative Folgen für die Existenz der Zelle haben. Somit geht eine Schädigung der äußeren Membran der Mitochondrien mit der Freisetzung von im Intermembranraum enthaltenen Substanzen, vor allem Cytochrom C und einigen anderen biologisch aktiven Substanzen, in das Zytoplasma einher, die Kettenreaktionen auslösen, die den programmierten Zelltod – Apoptose – verursachen. Durch die Schädigung der mitochondrialen DNA verzerren Reaktionen freier Radikale die genetische Information, die für die Bildung bestimmter Enzyme der Atmungskette erforderlich ist, die speziell in Mitochondrien produziert werden. Dies führt zu einer noch stärkeren Störung oxidativer Prozesse. Im Allgemeinen ist der genetische Apparat der Mitochondrien im Vergleich zum genetischen Apparat des Zellkerns weniger vor schädlichen Einflüssen geschützt, die die darin kodierte genetische Information verändern können. Infolgedessen kommt es während des gesamten Lebens zu Funktionsstörungen der Mitochondrien, beispielsweise während des Alterungsprozesses, bei der bösartigen Transformation der Zelle sowie vor dem Hintergrund erblicher mitochondrialer Erkrankungen, die mit einer Mutation der mitochondrialen DNA in der Eizelle einhergehen. Derzeit sind über 50 mitochondriale Mutationen beschrieben, die erblich bedingte degenerative Erkrankungen des Nerven- und Muskelsystems verursachen. Sie werden ausschließlich von der Mutter auf das Kind übertragen, da die Mitochondrien der Spermien nicht Teil der Zygote und damit des neuen Organismus sind.
Verletzung der Erhaltung und Übertragung genetischer Informationen
Der Zellkern enthält die meisten genetischen Informationen und sorgt so für seine normale Funktion. Durch selektive Genexpression koordiniert es die Zellaktivität während der Interphase, speichert genetische Informationen und erstellt und überträgt genetisches Material während der Zellteilung. DNA-Replikation und RNA-Transkription finden im Zellkern statt. Verschiedene pathogene Faktoren wie ultraviolette und ionisierende Strahlung, Oxidation durch freie Radikale, Chemikalien und Viren können die DNA schädigen. Es wird geschätzt, dass jede Zelle eines Warmblüters 1 Tag benötigt. verliert mehr als 10.000 Basen. Hier sollten wir Verstöße beim Kopieren während der Division hinzufügen. Wenn diese Schäden bestehen blieben, wäre die Zelle nicht überlebensfähig. Der Schutz liegt in der Existenz leistungsstarker Reparatursysteme wie UV-Endonuklease, Reparaturreplikations- und Rekombinationsreparatursystemen, die DNA-Schäden ersetzen. Genetische Defekte in Reparatursystemen führen zur Entstehung von Krankheiten, die durch eine erhöhte Empfindlichkeit gegenüber DNA-schädigenden Faktoren verursacht werden. Dabei handelt es sich um Xeroderma pigmentosum sowie um einige beschleunigte Alterungssyndrome, die mit einer erhöhten Neigung zur Entwicklung bösartiger Tumoren einhergehen.
Das System zur Regulierung der Prozesse der DNA-Replikation, der Transkription von Messenger-RNA (mRNA) und der Übersetzung genetischer Informationen aus Nukleinsäuren in die Struktur von Proteinen ist recht komplex und mehrstufig. Neben den Regulationskaskaden, die die Wirkung von Transkriptionsfaktoren mit einer Gesamtzahl von über 3000 auslösen, die bestimmte Gene aktivieren, gibt es auch ein mehrstufiges Regulationssystem, das durch kleine RNA-Moleküle vermittelt wird (interferierende RNA; RNAi). Das menschliche Genom, das aus etwa 3 Milliarden Purin- und Pyrimidinbasen besteht, enthält nur 2 % der Strukturgene, die für die Proteinsynthese verantwortlich sind. Der Rest sorgt für die Synthese regulatorischer RNAs, die gleichzeitig mit Transkriptionsfaktoren die Arbeit von Strukturgenen auf DNA-Ebene in Chromosomen aktivieren oder blockieren oder die Translationsprozesse von Messenger-RNA (mRNA) während der Bildung eines Polypeptidmoleküls beeinflussen das Zytoplasma. Verletzungen der genetischen Information können sowohl auf der Ebene von Strukturgenen als auch auf der Ebene des regulatorischen Teils der DNA auftreten und sich entsprechend in Form verschiedener Erbkrankheiten äußern.
In letzter Zeit wird viel Aufmerksamkeit auf Veränderungen im genetischen Material gelenkt, die während der individuellen Entwicklung eines Organismus auftreten und mit der Hemmung oder Aktivierung bestimmter Abschnitte der DNA und Chromosomen aufgrund ihrer Methylierung, Acetylierung und Phosphorylierung verbunden sind. Diese Veränderungen bleiben über einen langen Zeitraum bestehen, manchmal während des gesamten Lebens des Organismus von der Embryogenese bis ins hohe Alter, und werden als epigenomische Vererbung bezeichnet.
Die Vermehrung von Zellen mit veränderter genetischer Information wird auch durch Systeme (Faktoren) verhindert, die den Mitosezyklus steuern. Sie interagieren mit Cyclin-abhängigen Proteinkinasen und ihren katalytischen Untereinheiten – Cyclinen – und blockieren den vollständigen Mitosezyklus der Zelle, indem sie die Teilung an der Grenze zwischen der präsynthetischen und der synthetischen Phase (G1/S-Block) stoppen, bis die DNA-Reparatur abgeschlossen ist. und wenn dies nicht möglich ist, initiieren sie programmierte Todeszellen. Zu diesen Faktoren gehört das p53-Gen, dessen Mutation zum Verlust der Kontrolle über die Proliferation transformierter Zellen führt; es wird bei fast 50 % der Krebserkrankungen beim Menschen beobachtet. Der zweite Kontrollpunkt des Mitosezyklus liegt an der G2/M-Grenze. Hier wird die korrekte Verteilung des Chromosomenmaterials zwischen Tochterzellen in der Mitose oder Meiose mithilfe einer Reihe von Mechanismen gesteuert, die die Zellspindel, das Zentrum und die Zentromere (Kinetochoren) steuern. Die Unwirksamkeit dieser Mechanismen führt zu einer Störung der Verteilung der Chromosomen oder ihrer Teile, die sich durch das Fehlen eines Chromosoms in einer der Tochterzellen (Aneuploidie), das Vorhandensein eines zusätzlichen Chromosoms (Polyploidie) und die Trennung von a Teil eines Chromosoms (Deletion) und dessen Übertragung auf ein anderes Chromosom (Translokation). Solche Prozesse werden sehr häufig bei der Proliferation bösartig entarteter und transformierter Zellen beobachtet. Geschieht dies während der Meiose mit Keimzellen, führt dies entweder zum Tod des Fötus in einem frühen Stadium der Embryonalentwicklung oder zur Geburt eines Organismus mit einer Chromosomenerkrankung.
Eine unkontrollierte Zellproliferation während des Tumorwachstums entsteht als Folge von Mutationen in Genen, die die Zellproliferation steuern und als Onkogene bezeichnet werden. Von den mehr als 70 derzeit bekannten Onkogenen gehören die meisten zu Komponenten der Zellwachstumsregulation, einige sind Transkriptionsfaktoren, die die Genaktivität regulieren, sowie Faktoren, die die Zellteilung und das Zellwachstum hemmen. Ein weiterer Faktor, der die übermäßige Ausdehnung (Ausbreitung) proliferierender Zellen einschränkt, ist die Verkürzung der Chromosomenenden – Telomere, die sich aufgrund rein sterischer Wechselwirkungen nicht vollständig replizieren können, daher werden die Telomere nach jeder Zellteilung um verkürzt ein bestimmter Teil der Basen. So erschöpfen proliferierende Zellen eines erwachsenen Organismus nach einer bestimmten Anzahl von Teilungen (normalerweise 20 bis 100, abhängig von der Art des Organismus und seinem Alter) die Telomerlänge und die weitere Chromosomenreplikation stoppt. Dieses Phänomen tritt im Spermienepithel, in Enterozyten und in embryonalen Zellen aufgrund des Vorhandenseins des Enzyms Telomerase, das die Telomerlänge nach jeder Teilung wiederherstellt, nicht auf. In den meisten Zellen erwachsener Organismen ist die Telomerase blockiert, in Tumorzellen ist sie jedoch leider aktiviert.
Die Verbindung zwischen Kern und Zytoplasma sowie der Stofftransport in beide Richtungen erfolgt durch Poren in der Kernmembran unter Beteiligung spezieller, energieverbrauchender Transportsysteme. Auf diese Weise werden Energie und plastische Stoffe, Signalmoleküle (Transkriptionsfaktoren) zum Zellkern transportiert. Der Rückfluss transportiert mRNA- und Transfer-RNA-Moleküle (tRNA), Ribosomen, die für die Proteinsynthese in der Zelle notwendig sind, in das Zytoplasma. Der gleiche Stofftransportweg ist auch Viren, insbesondere HIV, eigen. Sie übertragen ihr genetisches Material in den Zellkern der Wirtszelle mit dessen weiterem Einbau in das Wirtsgenom und der Übertragung neu gebildeter viraler RNA in das Zytoplasma zur weiteren Synthese von Proteinen neuer Viruspartikel.
Verletzung von Syntheseprozessen
Proteinsyntheseprozesse finden in Zisternen des endoplasmatischen Retikulums statt, die eng mit Poren in der Kernmembran verbunden sind, durch die Ribosomen, tRNA und mRNA in das endoplasmatische Retikulum gelangen. Hier erfolgt die Synthese von Polypeptidketten, die anschließend im agranulären endoplasmatischen Retikulum und im Lamellenkomplex (Golgi-Komplex) ihre endgültige Form annehmen, wo sie eine posttranslationale Modifikation erfahren und sich mit Kohlenhydrat- und Lipidmolekülen verbinden. Neu gebildete Proteinmoleküle verbleiben nicht am Ort der Synthese, sondern durch einen komplexen regulierten Prozess namens Proteinkinese, werden aktiv in den isolierten Teil der Zelle übertragen, wo sie ihre beabsichtigte Funktion erfüllen. In diesem Fall ist ein sehr wichtiger Schritt die Strukturierung des übertragenen Moleküls in eine geeignete räumliche Konfiguration, die seine inhärente Funktion erfüllen kann. Diese Strukturierung erfolgt mit Hilfe spezieller Enzyme oder auf einer Matrix aus speziellen Proteinmolekülen – Chaperonen, die dem neu gebildeten oder durch äußere Einflüsse veränderten Proteinmolekül helfen, die richtige dreidimensionale Struktur zu erhalten. Im Falle einer schädlichen Wirkung auf die Zelle, wenn die Möglichkeit einer Störung der Struktur von Proteinmolekülen besteht (z. B. bei einem Anstieg der Körpertemperatur, einem Infektionsprozess, einer Vergiftung), steigt die Konzentration von Chaperonen in der Zelle scharf. Daher werden solche Moleküle auch genannt Stressproteine, oder Hitzeschockproteine. Eine Verletzung der Strukturierung eines Proteinmoleküls führt zur Bildung chemisch inerter Konglomerate, die sich bei Amyloidose, Alzheimer usw. in der Zelle oder außerhalb davon ablagern. Manchmal kann ein vorstrukturiertes ähnliches Molekül als Matrix dienen, und zwar in diesem Fall In diesem Fall sind bei einer fehlerhaften Primärstrukturierung auch alle nachfolgenden Moleküle fehlerhaft. Diese Situation tritt bei sogenannten Prionenerkrankungen (Scrapie beim Schaf, tollwütige Kuh, Kuru, Creutzfeldt-Jakob-Krankheit beim Menschen) auf, wenn ein Defekt in einem der Membranproteine einer Nervenzelle dazu führt, dass sich in der Zelle träge Massen ansammeln und Störung seiner lebenswichtigen Funktionen.
Eine Störung der Syntheseprozesse in einer Zelle kann in verschiedenen Stadien auftreten: RNA-Transkription im Zellkern, Translation von Polypeptiden in Ribosomen, posttranslationale Modifikation, Hypermethylierung und Glykosylierung des Beige-Moleküls, Transport und Verteilung von Proteinen in der Zelle und deren Entfernung nach draußen. In diesem Fall kann man eine Zunahme oder Abnahme der Anzahl der Ribosomen, den Abbau von Polyribosomen, eine Erweiterung der Zisternen des granulären endoplasmatischen Retikulums, einen Verlust von Ribosomen und die Bildung von Vesikeln und Vakuolen beobachten. Bei einer Vergiftung durch einen Blassen Haubentaucher wird daher das Enzym RNA-Polymerase beschädigt, was die Transkription stört. Durch die Inaktivierung des Elongationsfaktors stört Diphtherietoxin die Translationsprozesse und verursacht Myokardschäden. Die Ursache für eine Störung der Synthese einiger spezifischer Proteinmoleküle können Infektionserreger sein. Beispielsweise hemmen Herpesviren die Synthese und Expression von MHC-Antigenmolekülen, wodurch sie der Immunkontrolle teilweise entgehen können; Pestbazillen – die Synthese von Mediatoren akuter Entzündungen. Das Auftreten ungewöhnlicher Proteine kann deren weiteren Abbau stoppen und zur Ansammlung von inertem oder sogar toxischem Material führen. Dies kann bis zu einem gewissen Grad durch die Störung von Zerfallsprozessen erleichtert werden.
Störung von Zerfallsprozessen
Gleichzeitig mit der Proteinsynthese in der Zelle findet kontinuierlich dessen Abbau statt. Unter normalen Bedingungen hat dies wichtige regulatorische und formative Bedeutung, beispielsweise bei der Aktivierung inaktiver Formen von Enzymen, Proteinhormonen und Proteinen des Mitosezyklus. Normales Zellwachstum und normale Zellentwicklung erfordern ein fein kontrolliertes Gleichgewicht zwischen der Synthese und dem Abbau von Proteinen und Organellen. Im Prozess der Proteinsynthese wird jedoch aufgrund von Fehlern im Betrieb des Synthesegeräts, einer abnormalen Struktur des Proteinmoleküls und seiner Schädigung durch chemische und bakterielle Wirkstoffe ständig eine relativ große Anzahl fehlerhafter Moleküle gebildet. Schätzungen zufolge beträgt ihr Anteil etwa ein Drittel aller synthetisierten Proteine.
Säugetierzellen haben mehrere Hauptzellen Wege der Proteinzerstörung: durch lysosomale Proteasen (Pentidhydrolasen), kalziumabhängige Proteinasen (Endopeptidasen) und das Proteasomsystem. Darüber hinaus gibt es auch spezialisierte Proteinasen, beispielsweise Caspasen. Das Hauptorganell, in dem der Stoffabbau in eukaryotischen Zellen stattfindet, ist das Lysosom, das zahlreiche hydrolytische Enzyme enthält. Durch die Prozesse der Endozytose und verschiedener Arten der Autophagie in Lysosomen und Phagolysosomen werden sowohl defekte Proteinmoleküle als auch ganze Organellen zerstört: beschädigte Mitochondrien, Abschnitte der Plasmamembran, einige extrazelluläre Proteine und der Inhalt sekretorischer Granula.
Ein wichtiger Mechanismus für den Proteinabbau ist das Proteasom, eine multikatalytische Proteinasestruktur mit komplexer Struktur, die im Zytosol, im Zellkern, im endoplasmatischen Retikulum und auf der Zellmembran lokalisiert ist. Dieses Enzymsystem ist für den Abbau beschädigter Proteine sowie gesunder Proteine verantwortlich, die für eine normale Zellfunktion entfernt werden müssen. Dabei werden die zu zerstörenden Proteine vorab mit einem spezifischen Polypeptid, dem Ubiquitin, kombiniert. Allerdings können auch nicht-ubiquitinierte Proteine in Proteasomen teilweise zerstört werden. Der Abbau von Proteinmolekülen in Proteasomen in kurze Polypeptide (Prozessierung) und deren anschließende Präsentation zusammen mit Typ-I-MHC-Molekülen ist ein wichtiges Glied bei der Immunkontrolle der Antigenhomöostase im Körper. Wenn die Funktion des Proteasoms geschwächt ist, sammeln sich beschädigte und unnötige Proteine an, was mit der Zellalterung einhergeht. Eine Verletzung des Abbaus von Cyclin-abhängigen Proteinen führt zu einer Störung der Zellteilung, zum Abbau sekretorischer Proteine – zur Entwicklung einer Zystofibrose. Umgekehrt geht eine Erhöhung der Proteasomfunktion mit der Erschöpfung des Körpers einher (AIDS, Krebs).
Bei genetisch bedingten Störungen des Proteinabbaus ist der Organismus nicht lebensfähig und stirbt in den frühen Stadien der Embryogenese ab. Ist der Abbau von Fetten oder Kohlenhydraten gestört, kommt es zu Speicherkrankheiten (Thesaurismose). In diesem Fall reichert sich eine übermäßige Menge bestimmter Stoffe oder Produkte ihres unvollständigen Abbaus – Lipide, Polysaccharide – im Inneren der Zelle an, was die Funktion der Zelle erheblich beeinträchtigt. Am häufigsten wird dies in Leberepithelzellen (Hepatozyten), Neuronen, Fibroblasten und Makrophagozyten beobachtet.
Erworbene Störungen der Abbauprozesse von Stoffen können durch pathologische Prozesse (z. B. Eiweiß-, Fett-, Kohlenhydrat- und Pigmentabbau) entstehen und mit der Bildung ungewöhnlicher Stoffe einhergehen. Störungen im lysosomalen Proteolysesystem führen zu einer verminderten Anpassung während des Fastens oder zu erhöhtem Stress und zum Auftreten bestimmter endokriner Dysfunktionen – verminderte Insulin-, Thyreoglobulin-, Zytokin- und deren Rezeptorspiegel. Ein gestörter Proteinabbau verlangsamt die Wundheilung, führt zur Entstehung von Arteriosklerose und beeinträchtigt die Immunantwort. Bei Hypoxie, Veränderungen des intrazellulären pH-Werts, Strahlenschäden, gekennzeichnet durch erhöhte Peroxidation von Membranlipiden, sowie unter dem Einfluss lysosomotroper Substanzen – bakterielle Endotoxine, Metaboliten toxischer Pilze (Sporofusarin), Siliziumoxidkristalle – die Stabilität der Lysosomenmembran Veränderungen werden aktivierte lysosomale Enzyme in das Zytoplasma freigesetzt, was zur Zerstörung von Zellstrukturen und ihrem Tod führt.
Die Verbindung des Organismus mit der Umwelt ist aus physikalisch-chemischer Sicht ein offenes System, also ein System, in dem biochemische Prozesse ablaufen. Die Ausgangsstoffe stammen aus der Umwelt, die ebenfalls kontinuierlich gebildeten Stoffe werden nach außen transportiert. Das Gleichgewicht zwischen Geschwindigkeit und Konzentration der Produkte multidirektionaler Reaktionen im Körper ist bedingt, imaginär, da die Aufnahme und Entfernung von Substanzen nicht aufhört. Die kontinuierliche Verbindung mit der Umwelt ermöglicht es uns, einen lebenden Organismus als offenes System zu betrachten.
Für alle lebenden Zellen ist die Sonne die Energiequelle. Pflanzenzellen gewinnen mit Hilfe von Chlorophyll Energie aus dem Sonnenlicht und nutzen sie für Assimilationsreaktionen während des Prozesses der Photosynthese. Zellen von Tieren, Pilzen und Bakterien nutzen die Sonnenenergie indirekt beim Abbau organischer Substanzen, die von Erdpflanzen synthetisiert werden.
Bei der Zellatmung werden einige Nährstoffe der Zelle abgebaut und stellen so die für verschiedene Arten der Zellaktivität notwendige Energie bereit. Dieser Prozess findet in Organellen statt, die Mitochondrien genannt werden. Mitochondrien bestehen aus zwei Membranen: der äußeren, die die Organelle vom Zytoplasma trennt, und der inneren, die zahlreiche Falten bildet. Das Hauptprodukt der Atmung ist ATP. Es verlässt die Mitochondrien und dient als Energiequelle für viele chemische Reaktionen im Zytoplasma und in der Zellmembran. Wenn für die Zellatmung Sauerstoff benötigt wird, spricht man von aerober Atmung, treten Reaktionen jedoch ohne Sauerstoff auf, spricht man von anaerober Atmung.
Für jede Art von Arbeit, die in einer Zelle verrichtet wird, wird Energie in einer einzigen Form genutzt – in Form von Energie aus den Phosphatbindungen von ATP. ATP ist eine leicht mobile Verbindung. Die Bildung von ATP erfolgt an der inneren Membran der Mitochondrien. ATP wird in allen Zellen während der Atmung aufgrund der Oxidationsenergie von Kohlenhydraten, Fetten und anderen organischen Substanzen synthetisiert. In grünen Pflanzenzellen wird die Hauptmenge an ATP durch Sonnenenergie in Chloroplasten synthetisiert. Bei der Photosynthese produzieren sie ein Vielfaches mehr ATP als Mitochondrien. ATP zerfällt unter Aufbrechen der Phosphor-Sauerstoff-Bindungen und unter Freisetzung von Energie. Dies geschieht unter Einwirkung des Enzyms ATPase bei der Hydrolyse von ATP – der Anlagerung von Wasser unter Abspaltung eines Phosphorsäuremoleküls. Dadurch wird ATP in ADP umgewandelt, und wenn zwei Moleküle Phosphorsäure abgespalten werden, dann in AMP. Die Eliminierungsreaktion jedes Grammsäuremoleküls geht mit der Freisetzung von 40 kJ einher. Dies ist eine sehr große Energieabgabe, weshalb die Phosphor-Sauerstoff-Bindungen von ATP üblicherweise als makroergistisch (hochenergetisch) bezeichnet werden.
Die Verwendung von ATP in plastischen Austauschreaktionen erfolgt durch Kopplung mit der ATP-Hydrolyse. Moleküle verschiedener Stoffe werden durch Anlagerung der bei der Hydrolyse freigesetzten Phosphorgruppe aus dem ATP-Molekül, also durch Phosphorylierung, mit Energie aufgeladen.
Die Besonderheit von Phosphatderivaten besteht darin, dass sie die Zelle nicht verlassen können, obwohl ihre „entladenen“ Formen die Membran ungehindert passieren können. Dadurch bleiben phosphorylierte Moleküle in der Zelle, bis sie in entsprechenden Reaktionen eingesetzt werden.
Der umgekehrte Prozess der Umwandlung von ADP in ATP erfolgt durch die Zugabe eines Phosphorsäuremoleküls zu ADP, wodurch Wasser freigesetzt und eine große Energiemenge absorbiert wird.
Somit ist ATP eine universelle und direkte Energiequelle für die Zellaktivität. Dadurch entsteht ein einziger zellulärer Energiepool, der es ermöglicht, ihn von einem Bereich der Zelle in einen anderen umzuverteilen und zu transportieren.
Die Übertragung der Phosphatgruppe spielt eine wichtige Rolle bei chemischen Reaktionen wie dem Aufbau von Makromolekülen aus Monomeren. Beispielsweise können Aminosäuren erst nach vorheriger Phosphorylierung zu Peptiden zusammengefasst werden. Mechanische Prozesse der Kontraktion oder Bewegung, der Transport einer gelösten Substanz gegen einen Konzentrationsgradienten und andere Prozesse erfordern den Verbrauch von in ATP gespeicherter Energie.
Der Prozess des Energiestoffwechsels lässt sich wie folgt darstellen. Hochmolekulare organische Substanzen im Zytoplasma werden enzymatisch durch Hydrolyse in einfachere Substanzen umgewandelt, aus denen sie bestehen: Proteine – in Aminosäuren, Poly- und Disaccharide – in Monosaccharide (+ Glucose), Fette in Glycerin und Fettsäuren. Es finden keine oxidativen Prozesse statt, es wird wenig Energie freigesetzt, die nicht genutzt wird und in thermische Form übergeht. Die meisten Zellen nutzen zunächst Kohlenhydrate. Polysaccharide (Stärke bei Pflanzen und Glykogen bei Tieren) werden zu Glucose hydrolysiert. Die Glukoseoxidation erfolgt in drei Phasen: Glykolyse, oxidative Decarboxylierung (Krebs-Zyklus – Zitronensäurezyklus) und oxidative Phosphorylierung (Atmungskette). Im Zytoplasma findet eine Glykolyse statt, bei der ein Molekül Glucose in zwei Moleküle Brenztraubensäure unter Freisetzung von zwei Molekülen ATP gespalten wird. In Abwesenheit von Sauerstoff wird Brenztraubensäure entweder in Ethanol (Fermentation) oder Milchsäure (anaerobe Atmung) umgewandelt.
Bei der Glykolyse in tierischen Zellen zerfällt das aus sechs Kohlenstoffatomen bestehende Glucosemolekül in zwei Milchsäuremoleküle. Dieser Prozess ist mehrstufig. Es wird nacheinander von 13 Enzymen ausgeführt. Bei der alkoholischen Gärung entstehen aus einem Glucosemolekül zwei Moleküle Ethanol und zwei Moleküle CO2.
Die Glykolyse ist eine gemeinsame Phase der anaeroben und aeroben Atmung; die beiden anderen finden nur unter aeroben Bedingungen statt. Der Prozess der sauerstofffreien Oxidation, bei dem nur ein Teil der Energie der Metaboliten freigesetzt und genutzt wird, ist für anaerobe Organismen endgültig. In Gegenwart von Sauerstoff gelangt Brenztraubensäure in die Mitochondrien, wo sie durch eine Reihe aufeinanderfolgender Reaktionen vollständig aerob zu H2O und CO2 oxidiert wird und gleichzeitig ADP zu ATP phosphoryliert wird. In diesem Fall werden zwei ATP-Moleküle durch Glykolyse, zwei durch den Krebs-Zyklus und 34 durch die Atmungskette produziert. Die Nettoausbeute für die vollständige Oxidation eines Glucosemoleküls zu H2O und CO2 beträgt 38 Moleküle.
So erfolgt in aeroben Organismen der endgültige Abbau organischer Stoffe durch deren Oxidation mit Luftsauerstoff zu einfachen anorganischen Stoffen: CO2 und H2O. Dieser Prozess findet an den Kristallen der Mitochondrien statt. In diesem Fall wird die maximale Menge an freier Energie freigesetzt, von der ein erheblicher Teil in ATP-Molekülen reserviert ist. Es ist leicht zu erkennen, dass die aerobe Oxidation der Zelle den größten Teil der freien Energie liefert.
Durch den Katabolismus reichern sich energiereiche ATP-Moleküle in der Zelle an und CO2 und überschüssiges Wasser werden an die äußere Umgebung abgegeben.
Zuckermoleküle, die für die Atmung nicht benötigt werden, können in der Zelle gespeichert werden. Überschüssige Lipide werden entweder abgebaut, woraufhin die Abbauprodukte als Substrat für die Atmung in die Mitochondrien gelangen oder als Reserven in Form von Fetttröpfchen im Zytoplasma abgelagert werden. Proteine werden aus Aminosäuren aufgebaut, die in die Zelle gelangen. Die Proteinsynthese findet in Organellen statt, die Ribosomen genannt werden. Jedes Ribosom besteht aus zwei Unterpartikeln – einem großen und einem kleinen: Beide Unterpartikel umfassen Proteinmoleküle und RNA-Moleküle.
Ribosomen sind oft an ein spezielles Membransystem aus Zisternen und Vesikeln gebunden – das sogenannte endoplasmatische Retikulum (ER); In Zellen, die viel Protein produzieren, ist das endoplasmatische Retikulum oft sehr gut entwickelt und mit Ribosomen bedeckt. Manche Enzyme sind nur wirksam, wenn sie an eine Membran gebunden sind. Hier befinden sich die meisten Enzyme, die an der Lipidsynthese beteiligt sind. Somit ist das endoplasmatische Retikulum eine Art Zellwerkbank.
Darüber hinaus unterteilt das ER das Zytoplasma in separate Kompartimente, d. h. es trennt verschiedene chemische Prozesse, die gleichzeitig im Zytoplasma ablaufen, und verringert dadurch die Wahrscheinlichkeit, dass diese Prozesse einander stören.
Produkte, die von einer bestimmten Zelle produziert werden, werden häufig außerhalb der Zelle verwendet. In solchen Fällen passieren an Ribosomen synthetisierte Proteine die Membranen des endoplasmatischen Retikulums und werden in Membranvesikeln verpackt, die sich um sie herum bilden, die dann vom ER abgetrennt werden. Diese abgeflachten und übereinander gestapelten Vesikel bilden wie gestapelte Pfannkuchen eine charakteristische Struktur, die als Golgi-Komplex oder Golgi-Apparat bezeichnet wird. Während ihres Aufenthalts im Golgi-Apparat unterliegen Proteine bestimmten Veränderungen. Wenn es an der Zeit ist, die Zelle zu verlassen, verschmelzen die Membranvesikel mit der Zellmembran und entleeren sich, wobei ihr Inhalt ausgeschüttet wird, d. h. die Sekretion erfolgt durch Exozytose.
Der Golgi-Apparat produziert auch Lysosomen – Membranbeutel, die Verdauungsenzyme enthalten. Herauszufinden, wie eine Zelle bestimmte Proteine herstellt, verpackt und exportiert und wie sie „weiß“, welche Proteine sie für sich behalten sollte, ist einer der faszinierendsten Zweige der modernen Zytologie.
Die Membranen jeder Zelle sind ständig in Bewegung und verändern sich. ER-Membranen bewegen sich langsam durch die Zelle. Einzelne Abschnitte dieser Membranen trennen sich und bilden Vesikel, die vorübergehend Teil des Golgi-Apparats werden und dann durch den Prozess der Exozytose mit der Zellmembran verschmelzen.
Später wird das Membranmaterial in das Zytoplasma zurückgeführt und dort erneut verwendet.